

Trehalose 6-phosphate phosphatase is required for cell wall integrity and fungal virulence but not trehalose biosynthesis in the human fungal pathogen Aspergillus fumigatus
- PMID: 20545865
- PMCID: PMC2954268
- DOI: 10.1111/j.1365-2958.2010.07254.x
Trehalose 6-phosphate phosphatase is required for cell wall integrity and fungal virulence but not trehalose biosynthesis in the human fungal pathogen Aspergillus fumigatus
Abstract
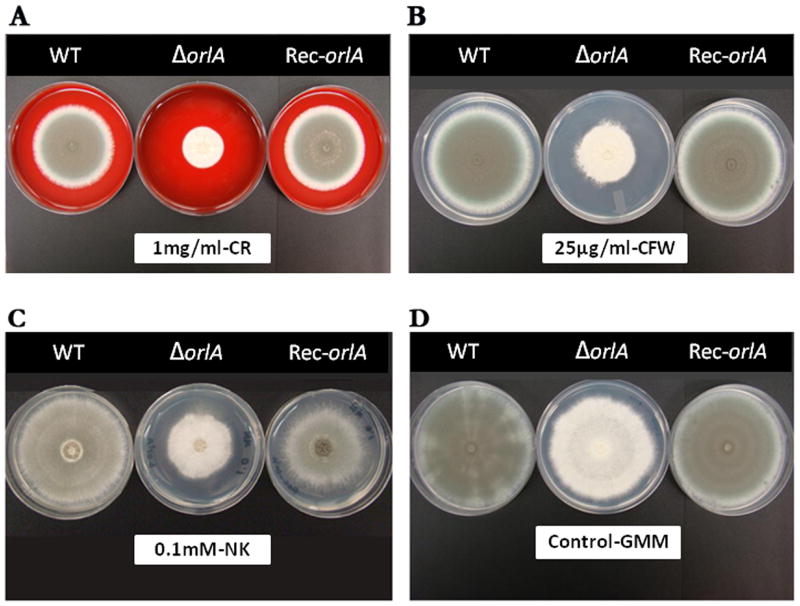
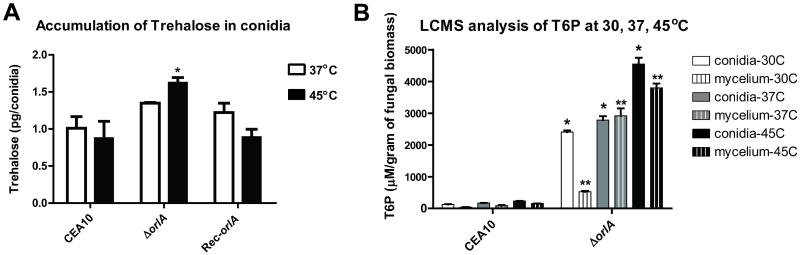
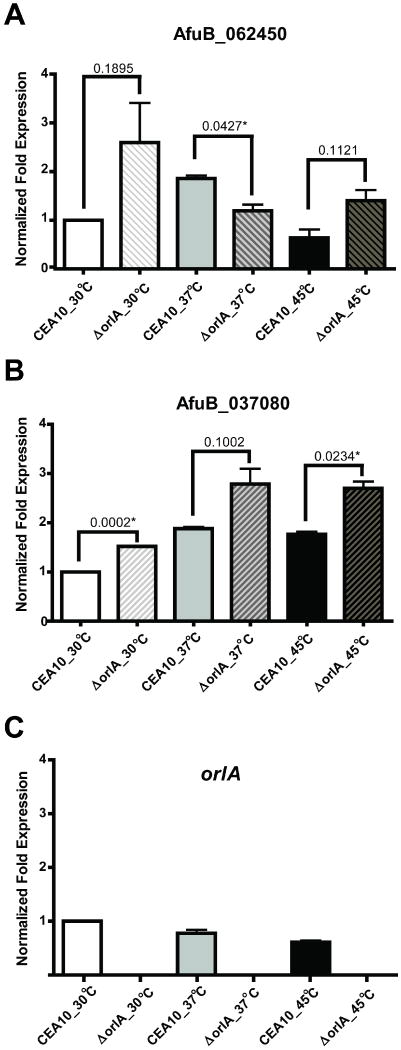
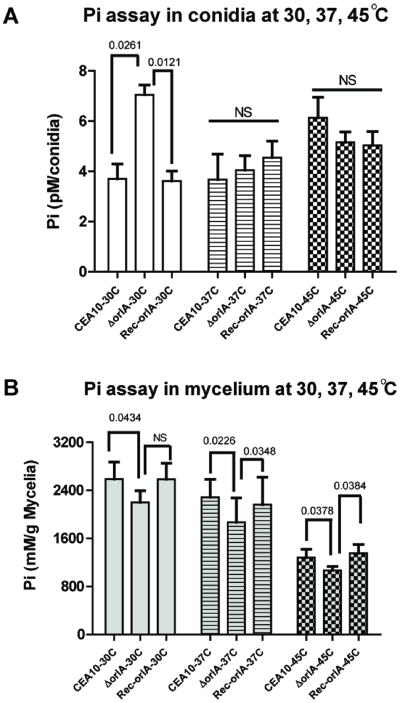
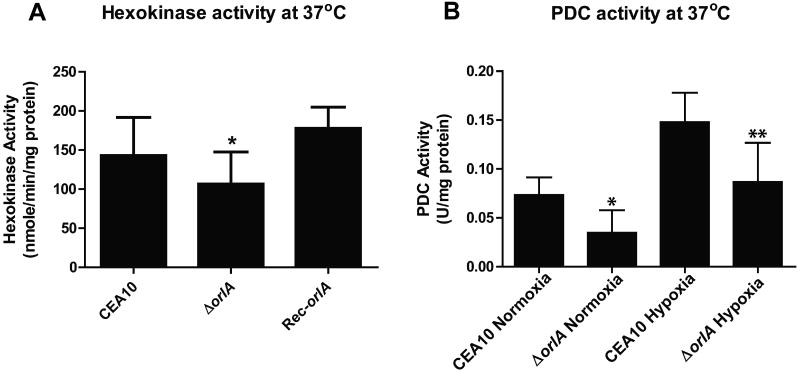
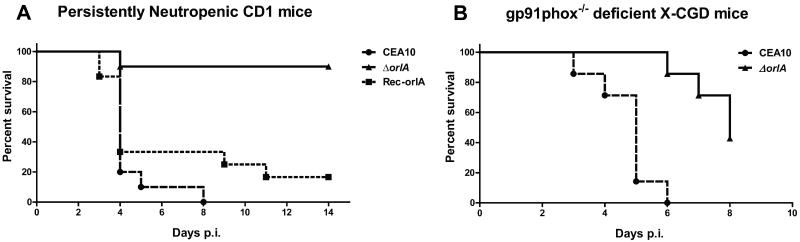
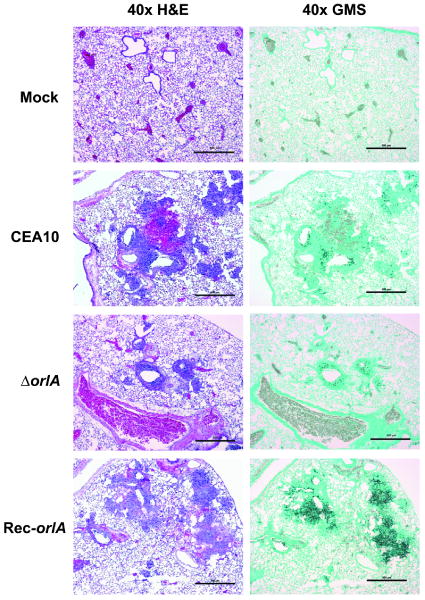
The trehalose biosynthesis pathway is critical for virulence in human and plant fungal pathogens. In this study, we tested the hypothesis that trehalose 6-phosphate phosphatase (T6PP) is required for Aspergillus fumigatus virulence. A mutant of the A. fumigatus T6PP, OrlA, displayed severe morphological defects related to asexual reproduction when grown on glucose (1%) minimal media. These defects could be rescued by addition of osmotic stabilizers, reduction in incubation temperature or increase in glucose levels (> 4%). Subsequent examination of the mutant with cell wall perturbing agents revealed a link between cell wall biosynthesis and trehalose 6-phosphate (T6P) levels. As expected, high levels of T6P accumulated in the absence of OrlA resulting in depletion of free inorganic phosphate and inhibition of hexokinase activity. Surprisingly, trehalose production persisted in the absence of OrlA. Further analyses revealed that A. fumigatus contains two trehalose phosphorylases that may be responsible for trehalose production in the absence of OrlA. Despite a normal growth rate under in vitro growth conditions, the orlA mutant was virtually avirulent in two distinct murine models of invasive pulmonary aspergillosis. Our results suggest that further study of this pathway will lead to new insights into regulation of fungal cell wall biosynthesis and virulence.
© 2010 Blackwell Publishing Ltd.
Figures
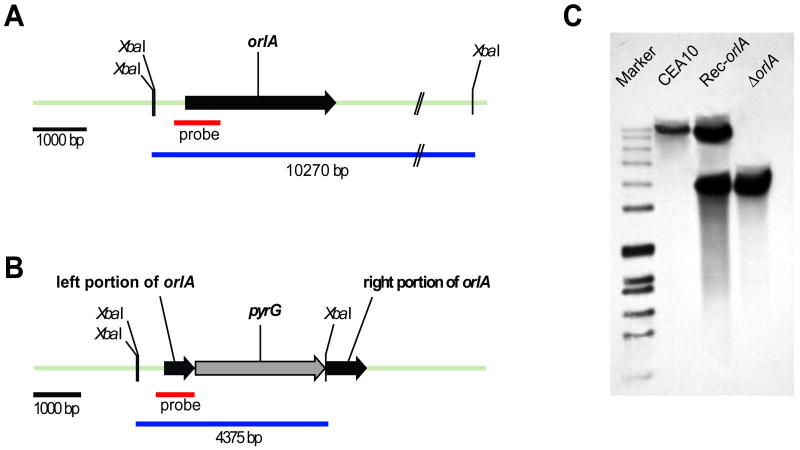
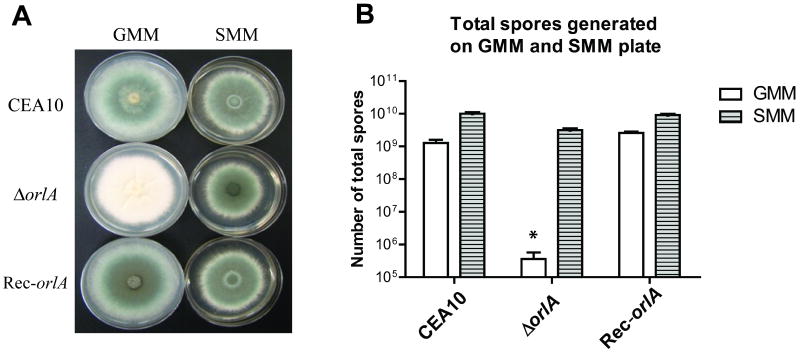
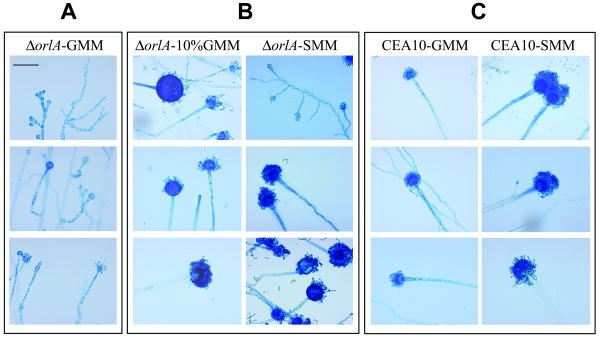
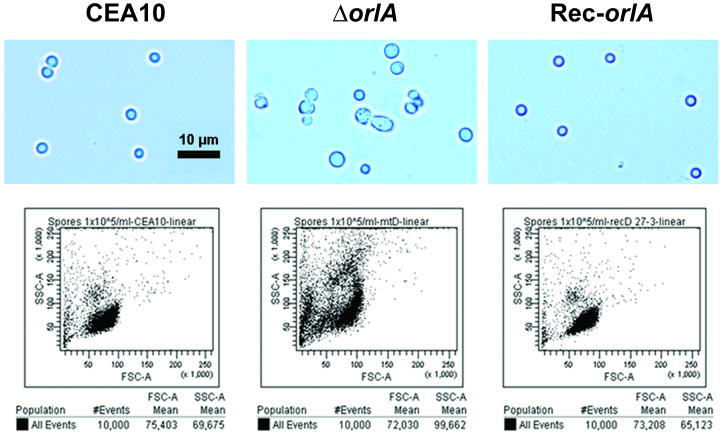
References
-
- Aisaka K, Masuda T. Production of trehalose phosphorylase by Catellatospora ferruginea. FEMS Microbiol Lett. 1995;131:47–51. - PubMed
-
- Aisaka K, Masuda T, Chikamune T, Kamitori K. Purification and characterization of trehalose phosphorylase from Catellatospora ferruginea. Biosci Biotechnol Biochem. 1998;62:782–787. - PubMed
-
- Anderson PJ, Karageuzian LN, Cheng HM, Epstein DL. Hexokinase of calf trabecular meshwork. Invest Ophthalmol Vis Sci. 1984;25:1258–1261. - PubMed
-
- Aufauvre-Brown A, Mellado E, Gow NAR, Holden DW. Aspergillus fumigatus chsE: A Gene Related to CHS3 of Saccharomyces cerevisiae and Important for Hyphal Growth and Conidiophore Development but Not Pathogenicity. Fungal Genet Biol. 1997;21:141–152. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
