

LPS-induced IL-8 activation in human intestinal epithelial cells is accompanied by specific histone H3 acetylation and methylation changes
- PMID: 20546607
- PMCID: PMC2893597
- DOI: 10.1186/1471-2180-10-172
LPS-induced IL-8 activation in human intestinal epithelial cells is accompanied by specific histone H3 acetylation and methylation changes
Abstract
Background: The release of LPS by bacteria stimulates both immune and specific epithelial cell types to release inflammatory mediators. It is known that LPS induces the release of IL-8 by intestinal mucosal cells. Because it is now emerging that bacteria may induce alteration of epigenetic patterns in host cells, we have investigated whether LPS-induced IL-8 activation in human intestinal epithelial cells involves changes of histone modifications and/or DNA methylation at IL-8 gene regulatory region.
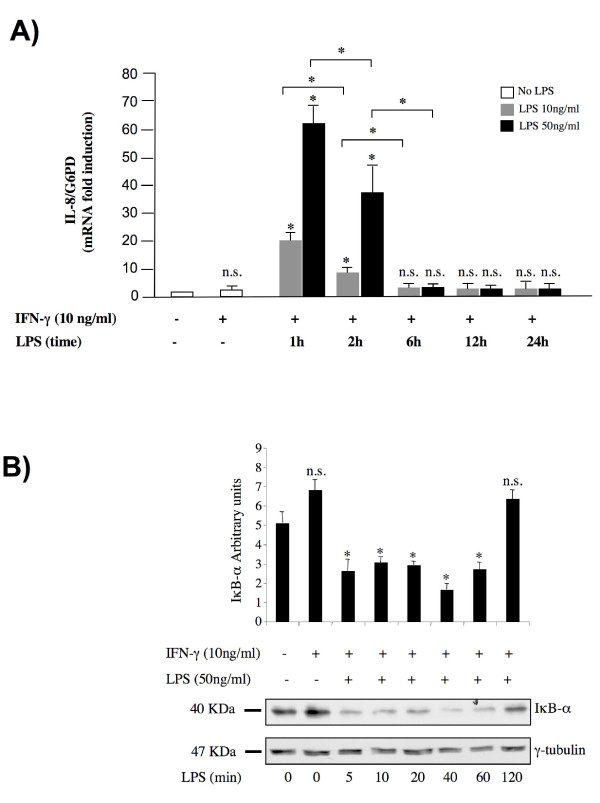
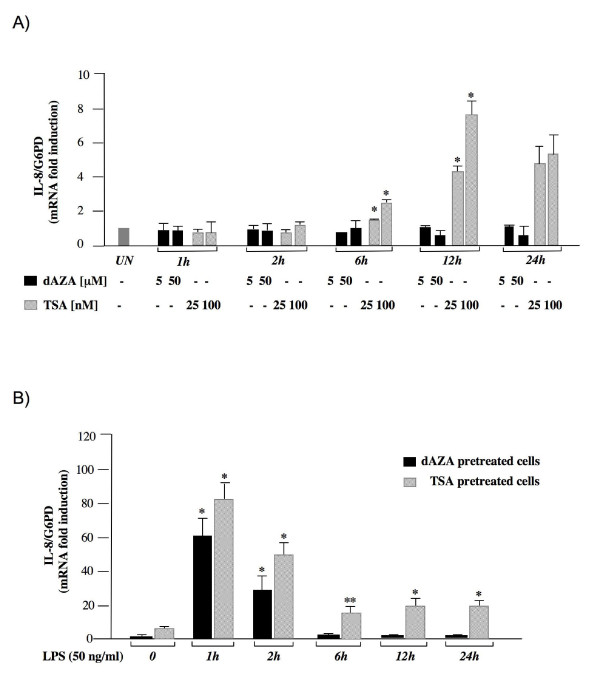
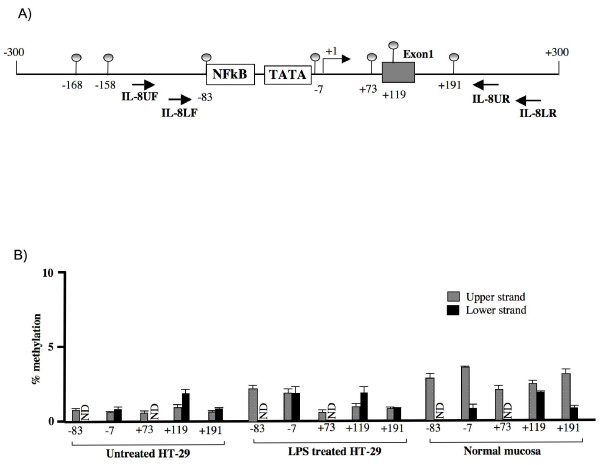
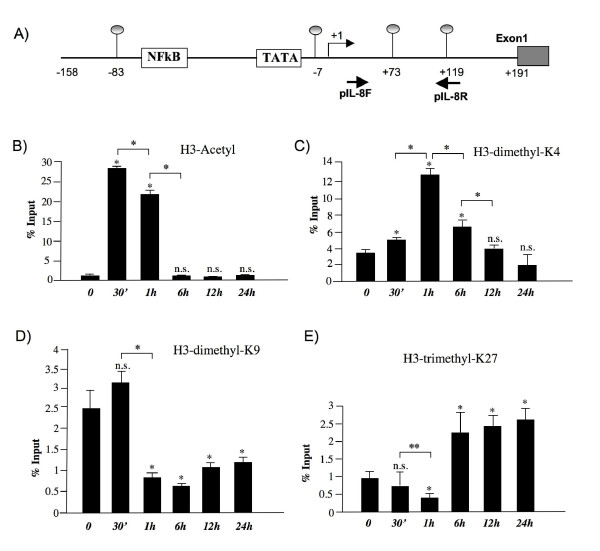
Results: RT-PCR analysis showed that IL-8 mRNA levels rapidly increase after exposure of HT-29 cells to LPS. DNA demethylating agents had no effects on IL-8 expression, suggesting that DNA methylation was not involved in IL-8 gene regulation. Consistently we found that 5 CpG sites located around IL-8 transcription start site (-83, -7, +73, +119, +191) were unmethylated on both lower and upper strand either in LPS treated or in untreated HT-29 cells, as well as in normal intestinal mucosa.Conversely, pretreatment of HT-29 cells with deacetylase inhibitors strengthened the LPS-mediated IL-8 activation. Inhibitors of histone deacetylases could induce IL-8 mRNA expression also in the absence of LPS, suggesting that chromatin modifications could be involved in IL-8 gene regulation. Chromatin immunoprecipitation analyses showed that, concurrently with IL-8 activation, transient specific changes in H3 acetylation and H3K4, H3K9 and H3K27 methylation occurred at IL-8 gene promoter during LPS stimulation. Changes of H3-acetyl, H3K4me2 and H3K9me2 levels occurred early, transiently and corresponded to transcriptional activity, while changes of H3K27me3 levels at IL-8 gene occurred later and were long lasting.
Conclusion: The results showed that specific chromatin modifications occurring at IL-8 gene, including histone H3 acetylation and methylation, mark LPS-mediated IL-8 activation in intestinal epithelial cells while it is unlikely that DNA methylation of IL-8 promoter is directly involved in IL-8 gene regulation in these cells.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
