

A critical cysteine is required for HMGB1 binding to Toll-like receptor 4 and activation of macrophage cytokine release
- PMID: 20547845
- PMCID: PMC2900689
- DOI: 10.1073/pnas.1003893107
A critical cysteine is required for HMGB1 binding to Toll-like receptor 4 and activation of macrophage cytokine release
Abstract
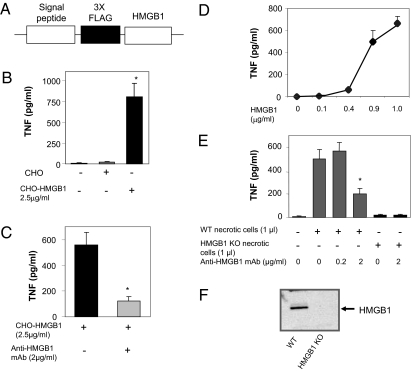
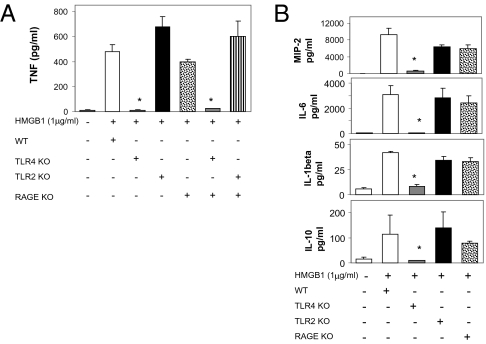
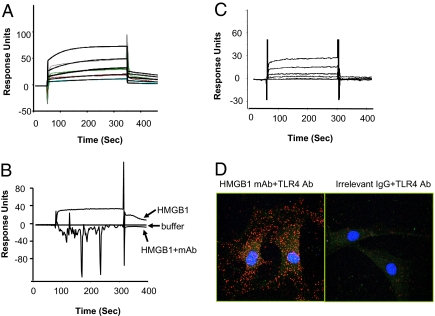
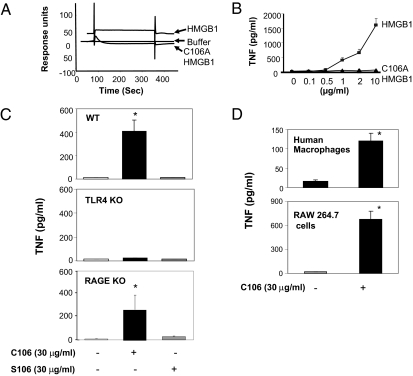
During infection, vertebrates develop "sickness syndrome," characterized by fever, anorexia, behavioral withdrawal, acute-phase protein responses, and inflammation. These pathophysiological responses are mediated by cytokines, including TNF and IL-1, released during the innate immune response to invasion. Even in the absence of infection, qualitatively similar physiological syndromes occur following sterile injury, ischemia reperfusion, crush injury, and autoimmune-mediated tissue damage. Recent advances implicate high-mobility group box 1 (HMGB1), a nuclear protein with inflammatory cytokine activities, in stimulating cytokine release. HMGB1 is passively released during cell injury and necrosis, or actively secreted during immune cell activation, positioning it at the intersection of sterile and infection-associated inflammation. To date, eight candidate receptors have been implicated in mediating the biological responses to HMGB1, but the mechanism of HMGB1-dependent cytokine release is unknown. Here we show that Toll-like receptor 4 (TLR4), a pivotal receptor for activation of innate immunity and cytokine release, is required for HMGB1-dependent activation of macrophage TNF release. Surface plasmon resonance studies indicate that HMGB1 binds specifically to TLR4, and that this binding requires a cysteine in position 106. A wholly synthetic 20-mer peptide containing cysteine 106 from within the cytokine-stimulating B box mediates TLR4-dependent activation of macrophage TNF release. Inhibition of TLR4 binding with neutralizing anti-HMGB1 mAb or by mutating cysteine 106 prevents HMGB1 activation of cytokine release. These results have implications for rationale, design, and development of experimental therapeutics for use in sterile and infectious inflammation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Wang H, et al. HMG-1 as a late mediator of endotoxin lethality in mice. Science. 1999;285:248–251. - PubMed
-
- Erlandsson Harris H, Andersson U. Mini-review: The nuclear protein HMGB1 as a proinflammatory mediator. Eur J Immunol. 2004;34:1503–1512. - PubMed
-
- Lotze MT, Tracey KJ. High-mobility group box 1 protein (HMGB1): nuclear weapon in the immune arsenal. Nat Rev Immunol. 2005;5:331–342. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
