

Membrane potential-dependent inactivation of voltage-gated ion channels in alpha-cells inhibits glucagon secretion from human islets
- PMID: 20547976
- PMCID: PMC2927942
- DOI: 10.2337/db09-1505
Membrane potential-dependent inactivation of voltage-gated ion channels in alpha-cells inhibits glucagon secretion from human islets
Abstract
Objective: To document the properties of the voltage-gated ion channels in human pancreatic alpha-cells and their role in glucagon release.
Research design and methods: Glucagon release was measured from intact islets. [Ca(2+)](i) was recorded in cells showing spontaneous activity at 1 mmol/l glucose. Membrane currents and potential were measured by whole-cell patch-clamping in isolated alpha-cells identified by immunocytochemistry.
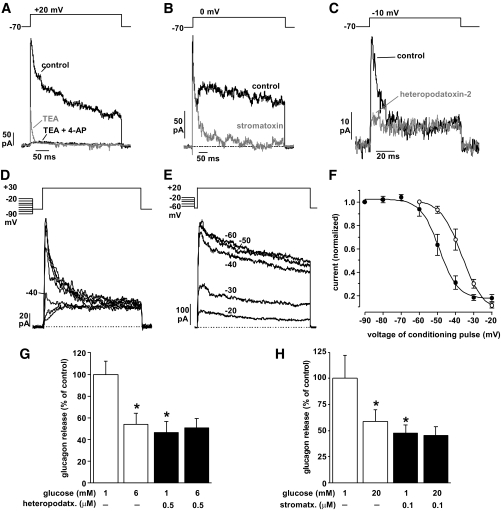
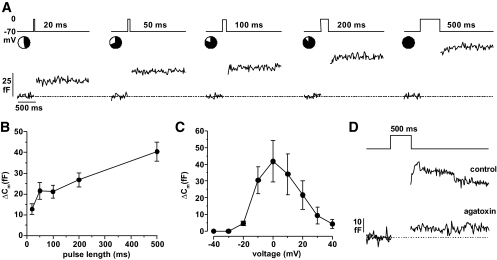
Result: Glucose inhibited glucagon secretion from human islets; maximal inhibition was observed at 6 mmol/l glucose. Glucagon secretion at 1 mmol/l glucose was inhibited by insulin but not by ZnCl(2). Glucose remained inhibitory in the presence of ZnCl(2) and after blockade of type-2 somatostatin receptors. Human alpha-cells are electrically active at 1 mmol/l glucose. Inhibition of K(ATP)-channels with tolbutamide depolarized alpha-cells by 10 mV and reduced the action potential amplitude. Human alpha-cells contain heteropodatoxin-sensitive A-type K(+)-channels, stromatoxin-sensitive delayed rectifying K(+)-channels, tetrodotoxin-sensitive Na(+)-currents, and low-threshold T-type, isradipine-sensitive L-type, and omega-agatoxin-sensitive P/Q-type Ca(2+)-channels. Glucagon secretion at 1 mmol/l glucose was inhibited by 40-70% by tetrodotoxin, heteropodatoxin-2, stromatoxin, omega-agatoxin, and isradipine. The [Ca(2+)](i) oscillations depend principally on Ca(2+)-influx via L-type Ca(2+)-channels. Capacitance measurements revealed a rapid (<50 ms) component of exocytosis. Exocytosis was negligible at voltages below -20 mV and peaked at 0 mV. Blocking P/Q-type Ca(2+)-currents abolished depolarization-evoked exocytosis.
Conclusions: Human alpha-cells are electrically excitable, and blockade of any ion channel involved in action potential depolarization or repolarization results in inhibition of glucagon secretion. We propose that voltage-dependent inactivation of these channels underlies the inhibition of glucagon secretion by tolbutamide and glucose.
Figures






References
-
- Cryer PE. Hypoglycaemia: the limiting factor in the glycaemic management of Type I and Type II diabetes. Diabetologia 2002;45:937–948 - PubMed
-
- Lefebvre PJ. (Ed.). Glucagon and Diabetes. In Handbook of Experimental Pharmacology. Berlin, Springer, 1996, p. 115–131
-
- Gromada J, Franklin I, Wollheim CB. Alpha-cells of the endocrine pancreas: 35 years of research but the enigma remains. Endocr Rev 2007;28:84–116 - PubMed
-
- Olofsson CS, Salehi A, Gopel SO, Holm C, Rorsman P. Palmitate stimulation of glucagon secretion in mouse pancreatic alpha-cells results from activation of L-type calcium channels and elevation of cytoplasmic calcium. Diabetes 2004;53:2836–2843 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
