

siRNA-mediated methylation of Arabidopsis telomeres
- PMID: 20548962
- PMCID: PMC2883606
- DOI: 10.1371/journal.pgen.1000986
siRNA-mediated methylation of Arabidopsis telomeres
Abstract
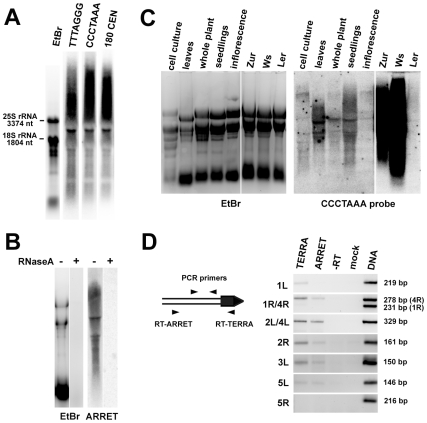
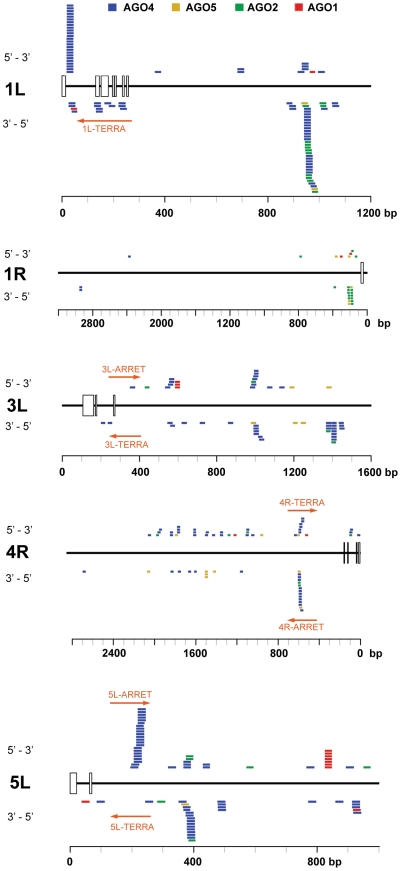
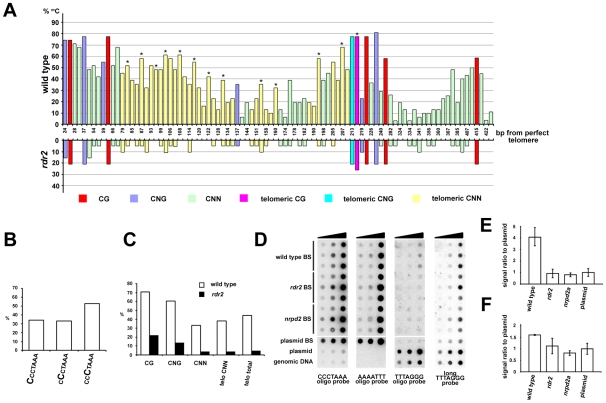
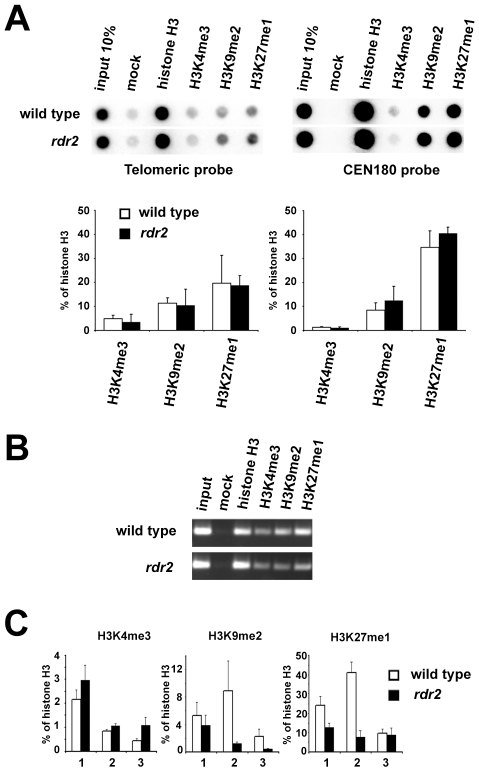
Chromosome termini form a specialized type of heterochromatin that is important for chromosome stability. The recent discovery of telomeric RNA transcripts in yeast and vertebrates raised the question of whether RNA-based mechanisms are involved in the formation of telomeric heterochromatin. In this study, we performed detailed analysis of chromatin structure and RNA transcription at chromosome termini in Arabidopsis. Arabidopsis telomeres display features of intermediate heterochromatin that does not extensively spread to subtelomeric regions which encode transcriptionally active genes. We also found telomeric repeat-containing transcripts arising from telomeres and centromeric loci, a portion of which are processed into small interfering RNAs. These telomeric siRNAs contribute to the maintenance of telomeric chromatin through promoting methylation of asymmetric cytosines in telomeric (CCCTAAA)(n) repeats. The formation of telomeric siRNAs and methylation of telomeres relies on the RNA-dependent DNA methylation pathway. The loss of telomeric DNA methylation in rdr2 mutants is accompanied by only a modest effect on histone heterochromatic marks, indicating that maintenance of telomeric heterochromatin in Arabidopsis is reinforced by several independent mechanisms. In conclusion, this study provides evidence for an siRNA-directed mechanism of chromatin maintenance at telomeres in Arabidopsis.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures



References
-
- Blasco MA. The epigenetic regulation of mammalian telomeres. Nat Rev Genet. 2007;8:299–309. - PubMed
-
- Ottaviani A, Gilson E, Magdinier F. Telomeric position effect: from the yeast paradigm to human pathologies? Biochimie. 2008;90:93–107. - PubMed
-
- Garcia-Cao M, O'Sullivan R, Peters AH, Jenuwein T, Blasco MA. Epigenetic regulation of telomere length in mammalian cells by the Suv39h1 and Suv39h2 histone methyltransferases. Nat Genet. 2004;36:94–99. - PubMed
-
- Gonzalo S, Garcia-Cao M, Fraga MF, Schotta G, Peters AH, et al. Role of the RB1 family in stabilizing histone methylation at constitutive heterochromatin. Nat Cell Biol. 2005;7:420–428. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
