

Regulation of vertebrate embryogenesis by the exon junction complex core component Eif4a3
- PMID: 20549732
- PMCID: PMC2922852
- DOI: 10.1002/dvdy.22330
Regulation of vertebrate embryogenesis by the exon junction complex core component Eif4a3
Abstract
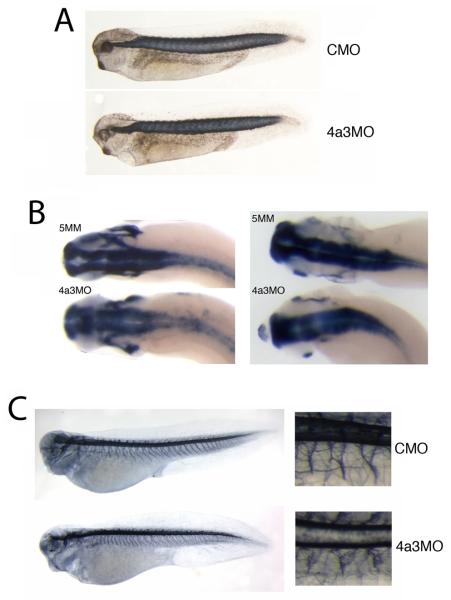
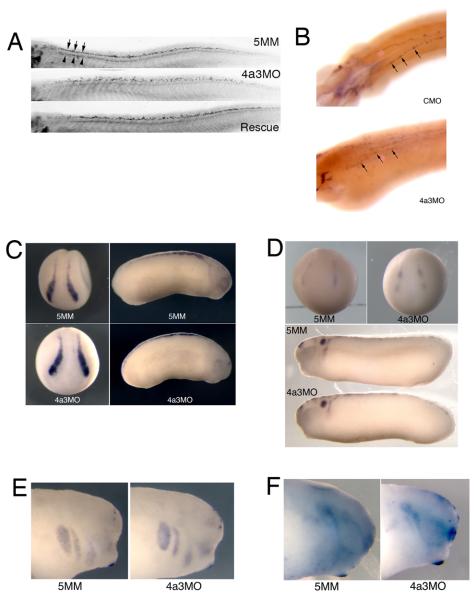
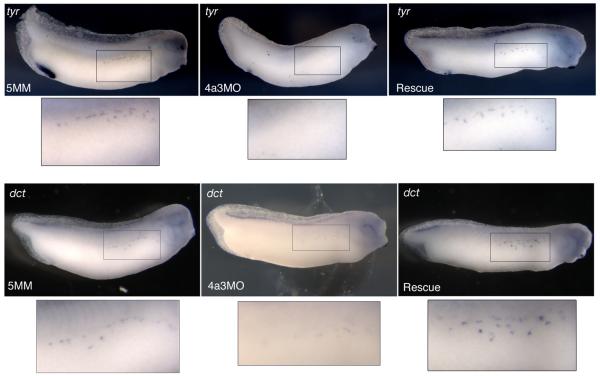
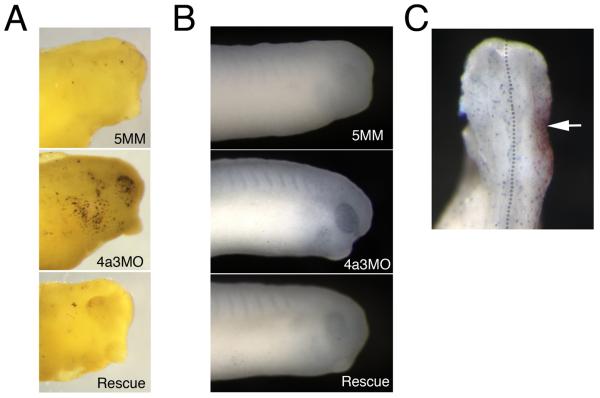
The establishment and maintenance of cellular identity are ultimately dependent upon the accurate regulation of gene expression, the process by which genetic information is used to synthesize functional gene products. The post-transcriptional, pre-translational regulation of RNA constitutes RNA processing, which plays a prominent role in the modulation of gene expression in differentiated animal cells. The multi-protein Exon Junction Complex (EJC) serves as a critical signaling hub within the network that underlies many RNA processing events. Here, we identify a requirement for the EJC during early vertebrate embryogenesis. Knockdown of the EJC component Eukaryotic initiation factor 4a3 (Eif4a3) in embryos of the frog Xenopus laevis results in full-body paralysis, with defects in sensory neuron, pigment cell, and cardiac development; similar phenotypes are seen following knockdown of other "core" EJC protein constituents. Our studies point to an essential role for the EJC in the development of neural plate border derivatives.
(c) 2010 Wiley-Liss, Inc.
Figures
Similar articles
-
Eif4a3 is required for accurate splicing of the Xenopus laevis ryanodine receptor pre-mRNA.Dev Biol. 2012 Dec 1;372(1):103-10. doi: 10.1016/j.ydbio.2012.08.013. Epub 2012 Aug 28. Dev Biol. 2012. PMID: 22944195 Free PMC article.
-
The Physiological Roles of the Exon Junction Complex in Development and Diseases.Cells. 2022 Apr 1;11(7):1192. doi: 10.3390/cells11071192. Cells. 2022. PMID: 35406756 Free PMC article. Review.
-
Two highly similar DEAD box proteins, OsRH2 and OsRH34, homologous to eukaryotic initiation factor 4AIII, play roles of the exon junction complex in regulating growth and development in rice.BMC Plant Biol. 2016 Apr 12;16:84. doi: 10.1186/s12870-016-0769-5. BMC Plant Biol. 2016. PMID: 27071313 Free PMC article.
-
Expression of the core exon-junction complex factor eukaryotic initiation factor 4A3 is increased during spatial exploration and striatally-mediated learning.Neuroscience. 2012 Dec 13;226:51-61. doi: 10.1016/j.neuroscience.2012.09.003. Epub 2012 Sep 12. Neuroscience. 2012. PMID: 22982623 Free PMC article.
-
Roles of the exon junction complex components in the central nervous system: a mini review.Rev Neurosci. 2018 Nov 27;29(8):817-824. doi: 10.1515/revneuro-2017-0113. Rev Neurosci. 2018. PMID: 29791316 Review.
Cited by
-
Thrombocytopenia-absent radius (TAR) syndrome due to compound inheritance for a 1q21.1 microdeletion and a low-frequency noncoding RBM8A SNP: a new familial case.Mol Cytogenet. 2015 Nov 5;8:87. doi: 10.1186/s13039-015-0188-6. eCollection 2015. Mol Cytogenet. 2015. PMID: 26550033 Free PMC article.
-
The exon junction complex is required for stem and progenitor cell maintenance in planarians.Dev Biol. 2020 Jan 1;457(1):119-127. doi: 10.1016/j.ydbio.2019.09.010. Epub 2019 Sep 23. Dev Biol. 2020. PMID: 31557470 Free PMC article.
-
Compound inheritance of a low-frequency regulatory SNP and a rare null mutation in exon-junction complex subunit RBM8A causes TAR syndrome.Nat Genet. 2012 Feb 26;44(4):435-9, S1-2. doi: 10.1038/ng.1083. Nat Genet. 2012. PMID: 22366785 Free PMC article.
-
Eif4a3 is required for accurate splicing of the Xenopus laevis ryanodine receptor pre-mRNA.Dev Biol. 2012 Dec 1;372(1):103-10. doi: 10.1016/j.ydbio.2012.08.013. Epub 2012 Aug 28. Dev Biol. 2012. PMID: 22944195 Free PMC article.
-
Zebrafish rbm8a and magoh mutants reveal EJC developmental functions and new 3'UTR intron-containing NMD targets.PLoS Genet. 2020 Jun 5;16(6):e1008830. doi: 10.1371/journal.pgen.1008830. eCollection 2020 Jun. PLoS Genet. 2020. PMID: 32502192 Free PMC article.
References
-
- Ballut L, Marchadier B, Baguet A, Tomasetto C, Seraphin B, Le Hir H. The exon junction core complex is locked onto RNA by inhibition of eIF4AIII ATPase activity. Nat Struct Mol Biol. 2005;12:861–869. - PubMed
-
- Chang YF, Imam JS, Wilkinson MF. The nonsense-mediated decay RNA surveillance pathway. Annu Rev Biochem. 2007;76:51–74. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
