

Millisecond timescale dynamics of human liver fatty acid binding protein: testing of its relevance to the ligand entry process
- PMID: 20550918
- PMCID: PMC2884256
- DOI: 10.1016/j.bpj.2010.03.047
Millisecond timescale dynamics of human liver fatty acid binding protein: testing of its relevance to the ligand entry process
Abstract
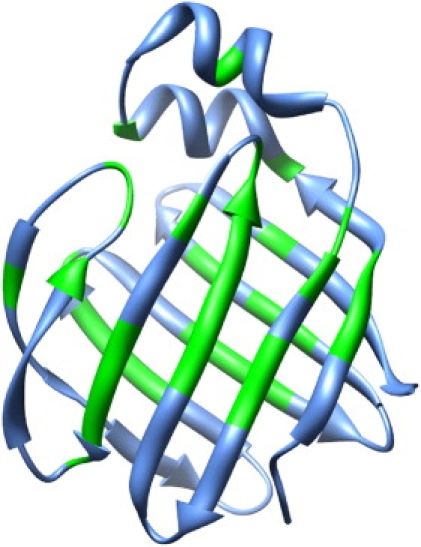
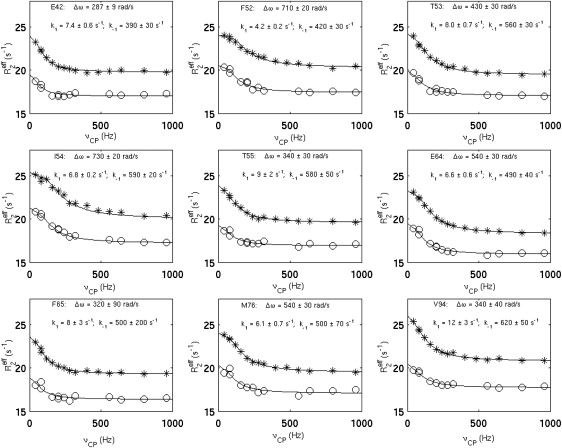
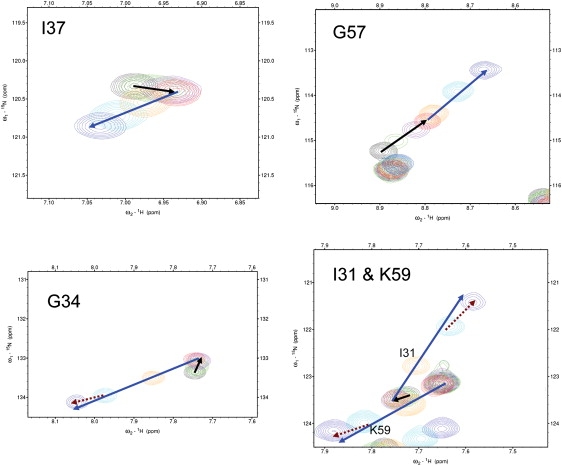
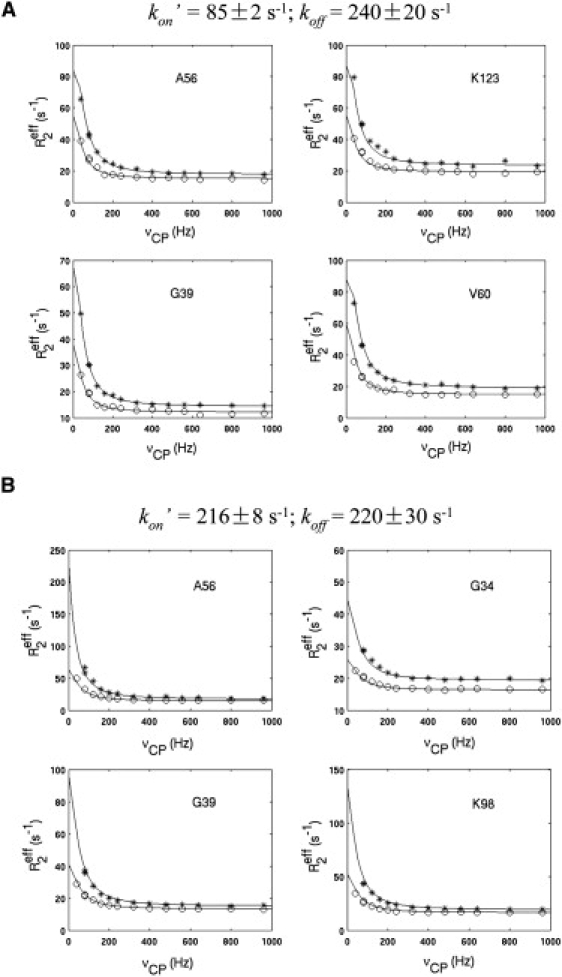
For over a decade, scientists have been attempting to know more about the conformational dynamics of fatty acid binding proteins (FABPs), to answer the puzzling question of how ligands could access the internalized binding site(s). Conformational exchange of FABPs on the microsecond to millisecond timescales has been found in many FABPs and offers an important hypothesis for the ligand entry mechanism. Despite the potential significance, the validity of this hypothesis has not been verified yet. In this study, the slow dynamics of human liver fatty acid binding protein (hLFABP) that was shown previously to be highly flexible on millisecond timescales was quantitatively characterized in detail. In addition, the interaction between hLFABP and 1,8-ANS was studied using NMR spectroscopy, and the kinetic rate of ANS association to hLFABP was measured. We believe the current result excludes the possibility that the intrinsic millisecond dynamics of hLFABP represents a critical conformational reorganization process required for ligand entry, but implies that it may represent the exchange between the apo-state and a state resembling the singly-bound conformation. Furthermore, we suggest these results show that the ligand-entry related functional dynamics could occur on the microsecond/submicrosecond timescales, highly encouraging future computational studies on this topic.
(c) 2010 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures
Similar articles
-
Buffer interference with protein dynamics: a case study on human liver fatty acid binding protein.Biophys J. 2009 Feb 18;96(4):1482-8. doi: 10.1016/j.bpj.2008.10.049. Biophys J. 2009. PMID: 19217864 Free PMC article.
-
Temperature dependence of backbone dynamics in human ileal bile acid-binding protein: implications for the mechanism of ligand binding.Biochemistry. 2014 Aug 12;53(31):5186-98. doi: 10.1021/bi500553f. Epub 2014 Jul 29. Biochemistry. 2014. PMID: 25073073
-
Solution-state molecular structure of apo and oleate-liganded liver fatty acid-binding protein.Biochemistry. 2007 Nov 6;46(44):12543-56. doi: 10.1021/bi701092r. Epub 2007 Oct 10. Biochemistry. 2007. PMID: 17927211
-
The role of dynamics in modulating ligand exchange in intracellular lipid binding proteins.Biochim Biophys Acta. 2014 Jul;1844(7):1268-78. doi: 10.1016/j.bbapap.2014.04.011. Epub 2014 Apr 21. Biochim Biophys Acta. 2014. PMID: 24768771 Review.
-
NMR methods for characterizing microsecond to millisecond dynamics in recognition and catalysis.Curr Opin Struct Biol. 2002 Oct;12(5):642-7. doi: 10.1016/s0959-440x(02)00369-x. Curr Opin Struct Biol. 2002. PMID: 12464317 Review.
Cited by
-
13Cα CEST experiment on uniformly 13C-labeled proteins.J Biomol NMR. 2015 Feb;61(2):89-94. doi: 10.1007/s10858-014-9888-1. Epub 2014 Dec 4. J Biomol NMR. 2015. PMID: 25465387
-
Rigidifying acyl carrier protein domain in iterative type I PKS CalE8 does not affect its function.Biophys J. 2012 Sep 5;103(5):1037-44. doi: 10.1016/j.bpj.2012.08.006. Biophys J. 2012. PMID: 23009853 Free PMC article.
-
Fatty Acid Binding Protein-1 (FABP1) and the Human FABP1 T94A Variant: Roles in the Endocannabinoid System and Dyslipidemias.Lipids. 2016 Jun;51(6):655-76. doi: 10.1007/s11745-016-4155-8. Epub 2016 Apr 27. Lipids. 2016. PMID: 27117865 Free PMC article. Review.
-
A combined computational-biophysical approach to understanding fatty acid binding to FABP7.Biophys J. 2023 Mar 7;122(5):741-752. doi: 10.1016/j.bpj.2023.02.003. Epub 2023 Feb 7. Biophys J. 2023. PMID: 36751130 Free PMC article.
-
Coexistence of multiple minor states of fatty acid binding protein and their functional relevance.Sci Rep. 2016 Sep 28;6:34171. doi: 10.1038/srep34171. Sci Rep. 2016. PMID: 27677899 Free PMC article.
References
-
- Storch J., Corsico B. The emerging functions and mechanisms of mammalian fatty acid-binding proteins. Annu. Rev. Nutr. 2008;28:73–95. - PubMed
-
- Thompson J., Reese-Wagoner A., Banaszak L. Liver fatty acid binding protein: species variation and the accommodation of different ligands. Biochim. Biophys. Acta. 1999;1441:117–130. - PubMed
-
- Xu Z., Bernlohr D.A., Banaszak L.J. The adipocyte lipid-binding protein at 1.6-A resolution. Crystal structures of the apoprotein and with bound saturated and unsaturated fatty acids. J. Biol. Chem. 1993;268:7874–7884. - PubMed
-
- Young A.C., Scapin G., Sacchettini J.C. Structural studies on human muscle fatty acid binding protein at 1.4 A resolution: binding interactions with three C18 fatty acids. Structure. 1994;2:523–534. - PubMed
-
- Lassen D., Lücke C., Rüterjans H. Three-dimensional structure of bovine heart fatty-acid-binding protein with bound palmitic acid, determined by multidimensional NMR spectroscopy. Eur. J. Biochem. 1995;230:266–280. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
