

ATM-dependent chromatin changes silence transcription in cis to DNA double-strand breaks
- PMID: 20550933
- PMCID: PMC2920610
- DOI: 10.1016/j.cell.2010.04.038
ATM-dependent chromatin changes silence transcription in cis to DNA double-strand breaks
Abstract
DNA double-strand breaks (DSBs) initiate extensive local and global alterations in chromatin structure, many of which depend on the ATM kinase. Histone H2A ubiquitylation (uH2A) on chromatin surrounding DSBs is one example, thought to be important for recruitment of repair proteins. uH2A is also implicated in transcriptional repression; an intriguing yet untested hypothesis is that this function is conserved in the context of DSBs. Using a novel reporter that allows for visualization of repair protein recruitment and local transcription in single cells, we describe an ATM-dependent transcriptional silencing program in cis to DSBs. ATM prevents RNA polymerase II elongation-dependent chromatin decondensation at regions distal to DSBs. Silencing is partially dependent on E3 ubiquitin ligases RNF8 and RNF168, whereas reversal of silencing relies on the uH2A deubiquitylating enzyme USP16. These findings give insight into the role of posttranslational modifications in mediating crosstalk between diverse processes occurring on chromatin.
Copyright 2010 Elsevier Inc. All rights reserved.
Figures
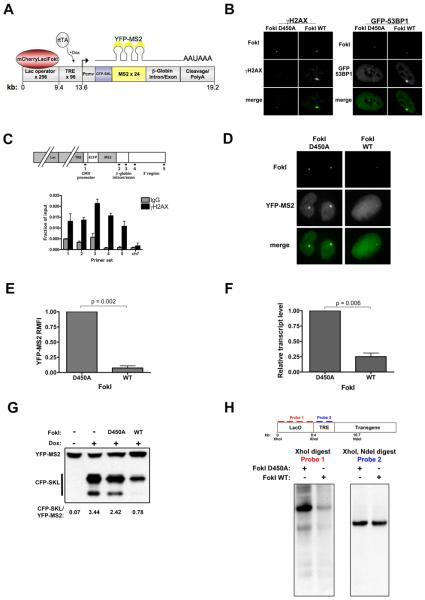
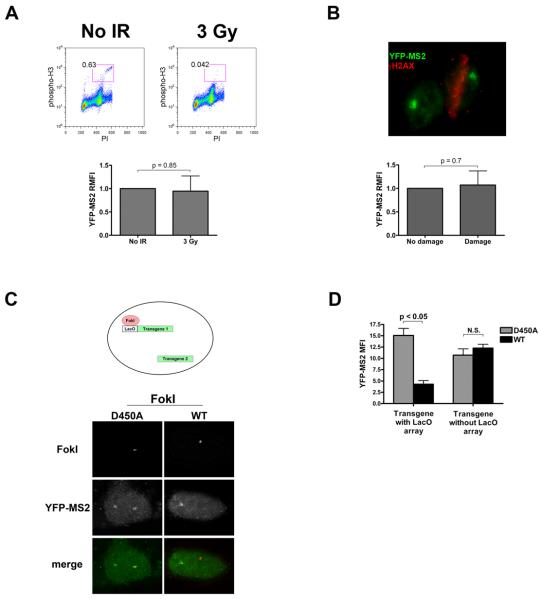
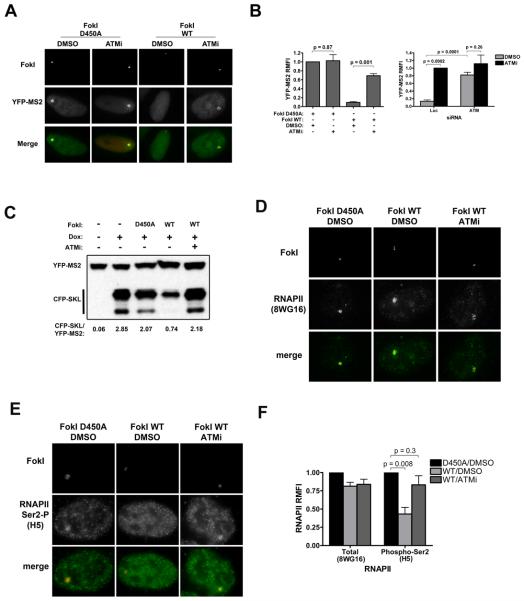
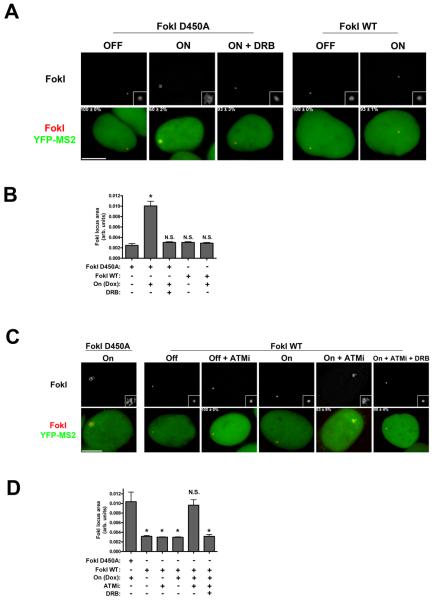
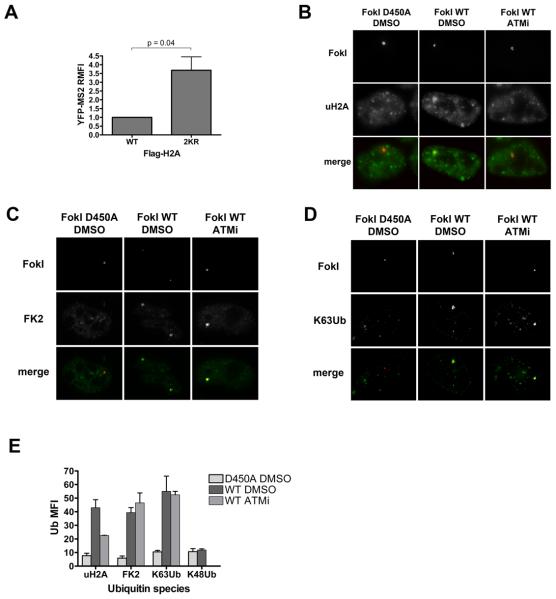
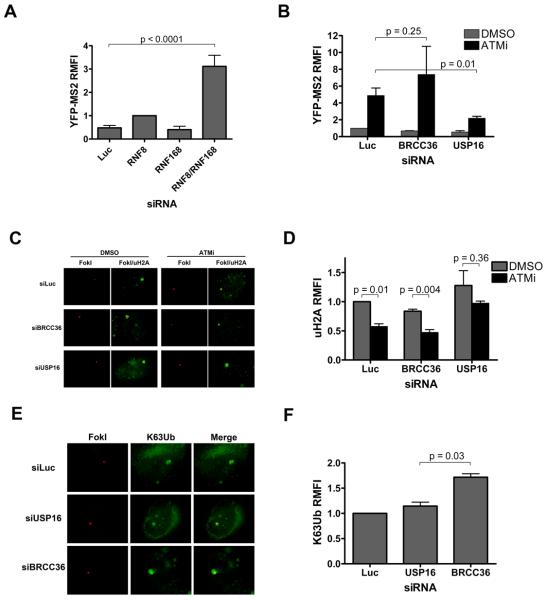
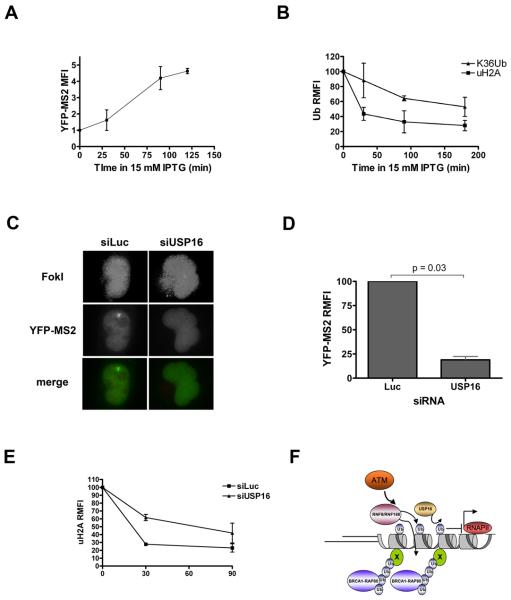
Comment in
-
ATM Creates a veil of transcriptional silence.Cell. 2010 Jun 11;141(6):924-6. doi: 10.1016/j.cell.2010.05.035. Cell. 2010. PMID: 20550929
References
-
- Anindya R, Aygun O, Svejstrup JQ. Damage-induced ubiquitylation of human RNA polymerase II by the ubiquitin ligase Nedd4, but not Cockayne syndrome proteins or BRCA1. Mol Cell. 2007;28:386–397. - PubMed
-
- Bartkova J, Horejsi Z, Koed K, Kramer A, Tort F, Zieger K, Guldberg P, Sehested M, Nesland JM, Lukas C, et al. DNA damage response as a candidate anti-cancer barrier in early human tumorigenesis. Nature. 2005;434:864–870. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
