

A novel phosphatase cascade regulates differentiation in Trypanosoma brucei via a glycosomal signaling pathway
- PMID: 20551176
- PMCID: PMC2885665
- DOI: 10.1101/gad.570310
A novel phosphatase cascade regulates differentiation in Trypanosoma brucei via a glycosomal signaling pathway
Abstract
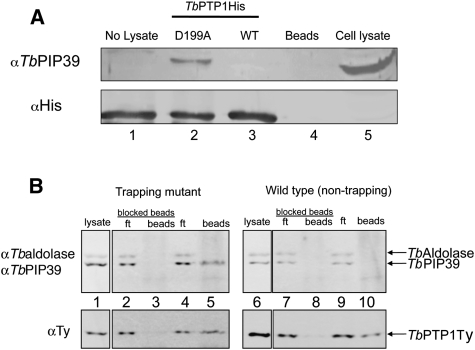
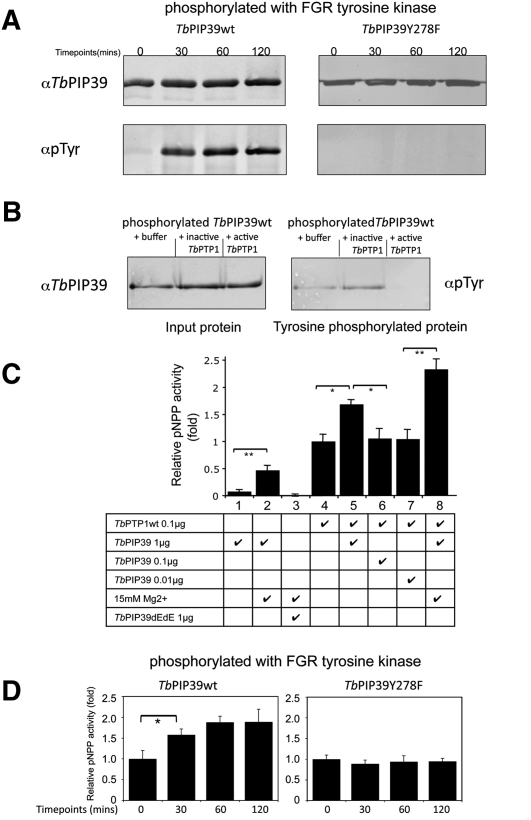
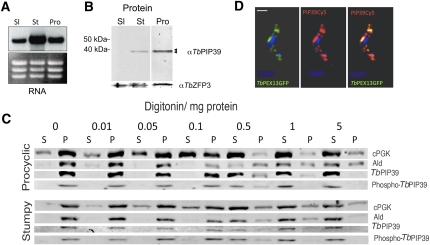
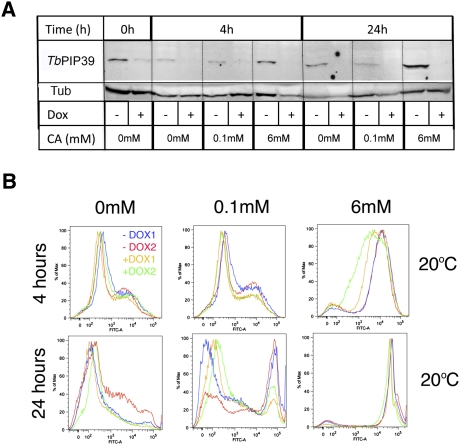
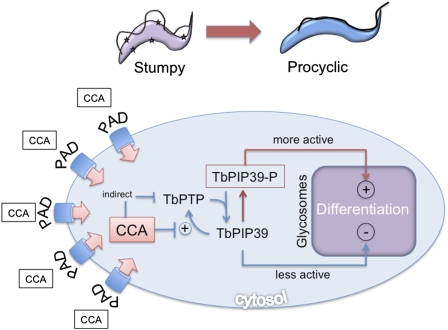
In the mammalian bloodstream, the sleeping sickness parasite Trypanosoma brucei is held poised for transmission by the activity of a tyrosine phosphatase, TbPTP1. This prevents differentiation of the transmissible "stumpy forms" until entry into the tsetse fly, whereupon TbPTP1 is inactivated and major changes in parasite physiology are initiated to allow colonization of the arthropod vector. Using a substrate-trapping approach, we identified the downstream step in this developmental signaling pathway as a DxDxT phosphatase, TbPIP39, which is activated upon tyrosine phosphorylation, and hence is negatively regulated by TbPTP1. In vitro, TbPIP39 promotes the activity of TbPTP1, thereby reinforcing its own repression, this being alleviated by the trypanosome differentiation triggers citrate and cis-aconitate, generating a potentially bistable regulatory switch. Supporting a role in signal transduction, TbPIP39 becomes rapidly tyrosine-phosphorylated during differentiation, and RNAi-mediated transcript ablation in stumpy forms inhibits parasite development. Interestingly, TbPIP39 localizes in glycosomes, peroxisome-like organelles that compartmentalize the trypanosome glycolytic reactions among other enzymatic activities. Our results invoke a phosphatase signaling cascade in which the developmental signal is trafficked to a unique metabolic organelle in the parasite: the glycosome. This is the first characterized environmental signaling pathway targeted directly to a peroxisome-like organelle in any eukaryotic cell.
Figures
References
-
- Albert MA, Haanstra JR, Hannaert V, Van Roy J, Opperdoes FR, Bakker BM, Michels PA 2005. Experimental and in silico analyses of glycolytic flux control in bloodstream form Trypanosoma brucei. J Biol Chem 280: 28306–28315 - PubMed
-
- Barrett MP, Burchmore RJ, Stich A, Lazzari JO, Frasch AC, Cazzulo JJ, Krishna S 2003. The trypanosomiases. Lancet 362: 1469–1480 - PubMed
-
- Bastin P, Bagherzadeh Z, Matthews KR, Gull K 1996. A novel epitope tag system to study protein targeting and organelle biogenesis in Trypanosoma brucei. Mol Biochem Parasitol 77: 235–239 - PubMed
-
- Biebinger S, Wirtz LE, Lorenz P, Clayton C 1997. Vectors for inducible expression of toxic gene products in bloodstream and procyclic Trypanosoma brucei. Mol Biochem Parasitol 85: 99–112 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases