

The nuclear protein Waharan is required for endosomal-lysosomal trafficking in Drosophila
- PMID: 20551180
- PMCID: PMC2894654
- DOI: 10.1242/jcs.060582
The nuclear protein Waharan is required for endosomal-lysosomal trafficking in Drosophila
Abstract
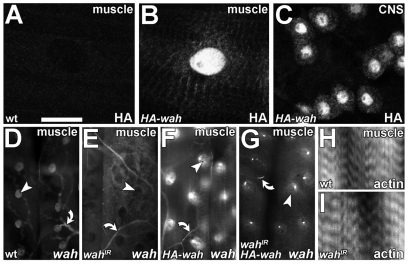
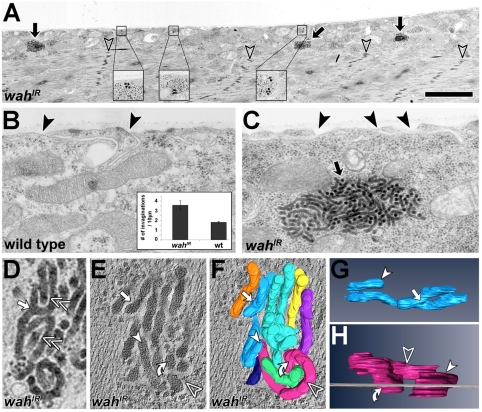
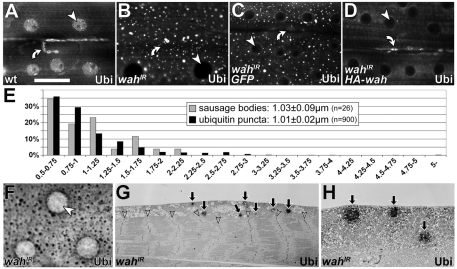
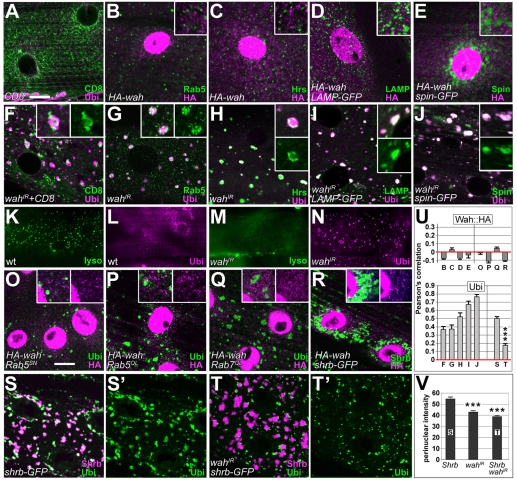
Here we report Drosophila Waharan (Wah), a 170-kD predominantly nuclear protein with two potential human homologues, as a newly identified regulator of endosomal trafficking. Wah is required for neuromuscular-junction development and muscle integrity. In muscles, knockdown of Wah caused novel accumulations of tightly packed electron-dense tubules, which we termed 'sausage bodies'. Our data suggest that sausage bodies coincide with sites at which ubiquitylated proteins and a number of endosomal and lysosomal markers co-accumulate. Furthermore, loss of Wah function generated loss of the acidic LysoTracker compartment. Together with data demonstrating that Wah acts earlier in the trafficking pathway than the Escrt-III component Drosophila Shrb (snf7 in Schizosaccharomyces pombe), our results indicate that Wah is essential for endocytic trafficking at the late endosome. Highly unexpected phenotypes result from Wah knockdown, in that the distribution of ubiquitylated cargos and endolysosomal morphologies are affected despite Wah being a predominant nuclear protein. This finding suggests the existence of a relationship between nuclear functions and endolysosomal trafficking. Future studies of Wah function will give us insights into this interesting phenomenon.
Figures
References
-
- Bate M. (1993). The mesoderm and its derivatives. In The development of Drosophila melanogaster, vol. 2 (eds Bate M., Martínez Arias A.), pp. 1013-1090 Cold Spring Harbor: CSH Laboratory Press;
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
