

A distinct type of glycerol-3-phosphate acyltransferase with sn-2 preference and phosphatase activity producing 2-monoacylglycerol
- PMID: 20551224
- PMCID: PMC2900678
- DOI: 10.1073/pnas.0914149107
A distinct type of glycerol-3-phosphate acyltransferase with sn-2 preference and phosphatase activity producing 2-monoacylglycerol
Abstract
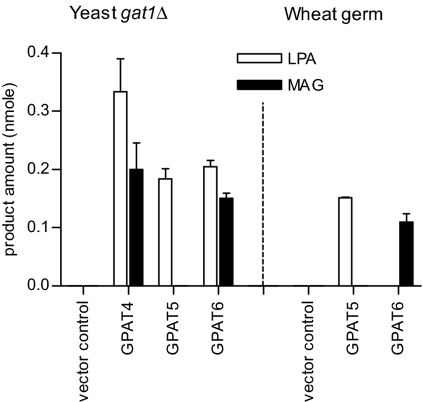
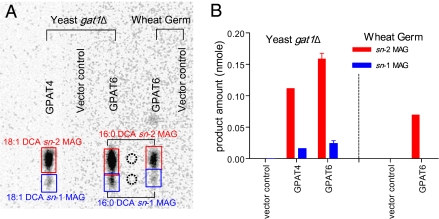
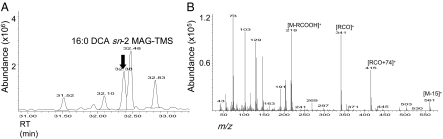
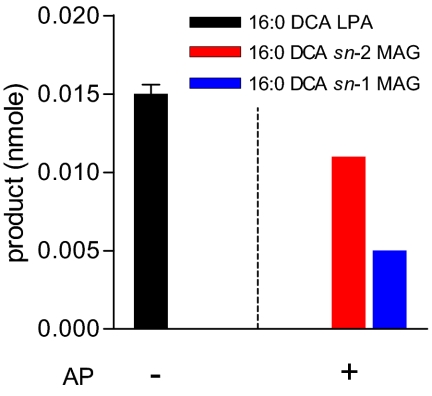
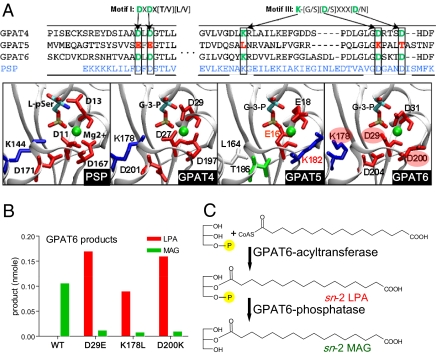
The first step in assembly of membrane and storage glycerolipids is acylation of glycerol-3-phosphate (G3P). All previously characterized membrane-bound, eukaryotic G3P acyltransferases (GPATs) acylate the sn-1 position to produce lysophosphatidic acid (1-acyl-LPA). Cutin is a glycerolipid with omega-oxidized fatty acids and glycerol as integral components. It occurs as an extracellular polyester on the aerial surface of all plants, provides a barrier to pathogens and resistance to stress, and maintains organ identity. We have determined that Arabidopsis acyltransferases GPAT4 and GPAT6 required for cutin biosynthesis esterify acyl groups predominantly to the sn-2 position of G3P. In addition, these acyltransferases possess a phosphatase domain that results in sn-2 monoacylglycerol (2-MAG) rather than LPA as the major product. Such bifunctional activity has not been previously described in any organism. The possible roles of 2-MAGs as intermediates in cutin synthesis are discussed. GPAT5, which is essential for the accumulation of suberin aliphatics, also exhibits a strong preference for sn-2 acylation. However, phosphatase activity is absent and 2-acyl-LPA is the major product. Clearly, plant GPATs can catalyze more reactions than the sn-1 acylation by which they are currently categorized. Close homologs of GPAT4-6 are present in all land plants, but not in animals, fungi or microorganisms (including algae). Thus, these distinctive acyltransferases may have been important for evolution of extracellular glycerolipid polymers and adaptation of plants to a terrestrial environment. These results provide insight into the biosynthetic assembly of cutin and suberin, the two most abundant glycerolipid polymers in nature.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Gimeno RE, Cao J. Mammalian glycerol-3-phosphate acyltransferases: New genes for an old activity. J Lipid Res. 2008;49:2079–2088. - PubMed
-
- Zheng Z, Zou J. The initial step of the glycerolipid pathway: Identification of glycerol 3-phosphate/dihydroxyacetone phosphate dual substrate acyltransferases in Saccharomyces cerevisiae. J Biol Chem. 2001;276:41710–41716. - PubMed
-
- Murata N, Tasaka Y. Glycerol-3-phosphate acyltransferase in plants. Biochim Biophys Acta. 1997;1348:10–16. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
