

The evolution of epitype
- PMID: 20551346
- PMCID: PMC2910975
- DOI: 10.1105/tpc.110.075481
The evolution of epitype
Abstract
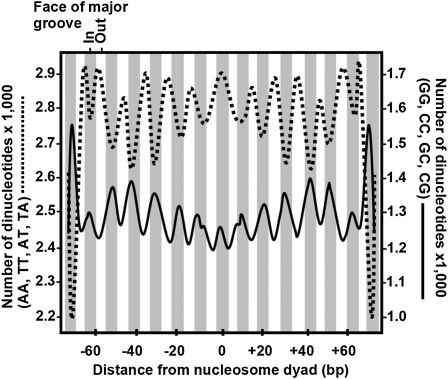
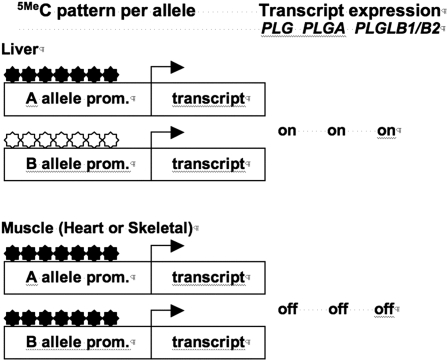
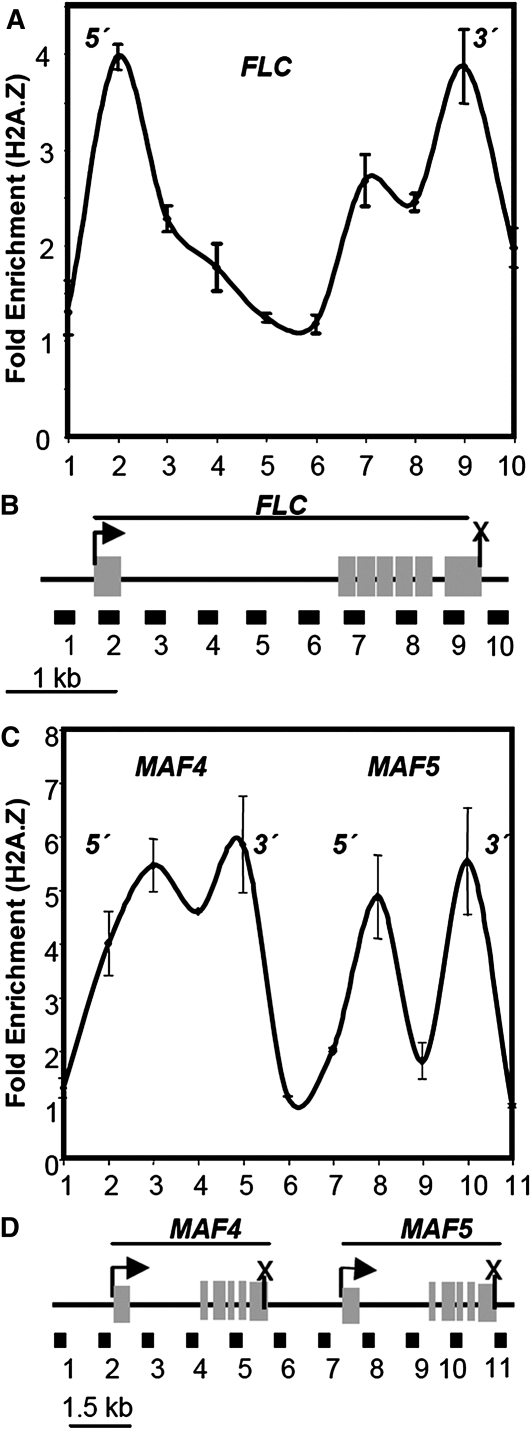
The epitype of a single gene or entire genome is determined by cis-linked differences in chromatin structure. I explore the hypothesis that "epitype and associated phenotypes evolve by gene duplication, divergence, and subfunctionalization" parallel to models for the evolution of genotype. This hypothesis is dissected by considering the relationship between epigenetic control and phenotype, the phylogenetic evidence that epitype evolves from ancestral genes following gene duplication, and the possible evolutionary rates of change for different epitypes. Initial supporting arguments for this hypothesis are discussed based on conserved patterns of nucleosome phasing, DNA methylation, and histone variant H2AZ deposition that appear to contribute to the inheritance of epitype in plants and animals. However, patterns of histone modification in recent segmental chromosome duplications are not well conserved. A continued experimental examination of the link between gene phylogeny and epitype and the evolution of epigenetically determined phenotypes is needed to further explore this hypothesis.
Figures
Similar articles
-
The influence of DNA sequence on epigenome-induced pathologies.Epigenetics Chromatin. 2012 Jul 20;5(1):11. doi: 10.1186/1756-8935-5-11. Epigenetics Chromatin. 2012. PMID: 22818522 Free PMC article.
-
Histone modification pattern evolution after yeast gene duplication.BMC Evol Biol. 2012 Jul 9;12:111. doi: 10.1186/1471-2148-12-111. BMC Evol Biol. 2012. PMID: 22776110 Free PMC article.
-
An integrated genomics analysis of epigenetic subtypes in human breast tumors links DNA methylation patterns to chromatin states in normal mammary cells.Breast Cancer Res. 2016 Feb 29;18(1):27. doi: 10.1186/s13058-016-0685-5. Breast Cancer Res. 2016. PMID: 26923702 Free PMC article.
-
Epigenomics in stress tolerance of plants under the climate change.Mol Biol Rep. 2023 Jul;50(7):6201-6216. doi: 10.1007/s11033-023-08539-6. Epub 2023 Jun 9. Mol Biol Rep. 2023. PMID: 37294468 Review.
-
Transgenerational inheritance: how impacts to the epigenetic and genetic information of parents affect offspring health.Hum Reprod Update. 2019 Sep 11;25(5):518-540. doi: 10.1093/humupd/dmz017. Hum Reprod Update. 2019. PMID: 31374565
Cited by
-
The histone modification H3K27me3 is retained after gene duplication and correlates with conserved noncoding sequences in Arabidopsis.Genome Biol Evol. 2014 Mar;6(3):572-9. doi: 10.1093/gbe/evu040. Genome Biol Evol. 2014. PMID: 24567304 Free PMC article.
-
Sustainable harvest: managing plasticity for resilient crops.Plant Biotechnol J. 2014 Jun;12(5):517-33. doi: 10.1111/pbi.12198. Plant Biotechnol J. 2014. PMID: 24891039 Free PMC article. Review.
-
Genetic variation and expression diversity between grain and sweet sorghum lines.BMC Genomics. 2013 Jan 16;14:18. doi: 10.1186/1471-2164-14-18. BMC Genomics. 2013. PMID: 23324212 Free PMC article.
-
The influence of DNA sequence on epigenome-induced pathologies.Epigenetics Chromatin. 2012 Jul 20;5(1):11. doi: 10.1186/1756-8935-5-11. Epigenetics Chromatin. 2012. PMID: 22818522 Free PMC article.
References
-
- Abbosh P.H., Montgomery J.S., Starkey J.A., Novotny M., Zuhowski E.G., Egorin M.J., Moseman A.P., Golas A., Brannon K.M., Balch C., Huang T.H., Nephew K.P. (2006). Dominant-negative histone H3 lysine 27 mutant derepresses silenced tumor suppressor genes and reverses the drug-resistant phenotype in cancer cells. Cancer Res. 66: 5582–5591 - PubMed
-
- Albert I., Mavrich T.N., Tomsho L.P., Qi J., Zanton S.J., Schuster S.C., Pugh B.F. (2007). Translational and rotational settings of H2A.Z nucleosomes across the Saccharomyces cerevisiae genome. Nature 446: 572–576 - PubMed
-
- Angermayr M., Oechsner U., Bandlow W. (2003). Reb1p-dependent DNA bending effects nucleosome positioning and constitutive transcription at the yeast profilin promoter. J. Biol. Chem. 278: 17918–17926 - PubMed
-
- Ay A., Arnosti D.N. (2010). Nucleosome positioning: An essential component of the enhancer regulatory code? Curr. Biol. 20: R404–R406 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
