

Detecting population structure in a high gene-flow species, Atlantic herring (Clupea harengus): direct, simultaneous evaluation of neutral vs putatively selected loci
- PMID: 20551979
- PMCID: PMC3183876
- DOI: 10.1038/hdy.2010.71
Detecting population structure in a high gene-flow species, Atlantic herring (Clupea harengus): direct, simultaneous evaluation of neutral vs putatively selected loci
Abstract
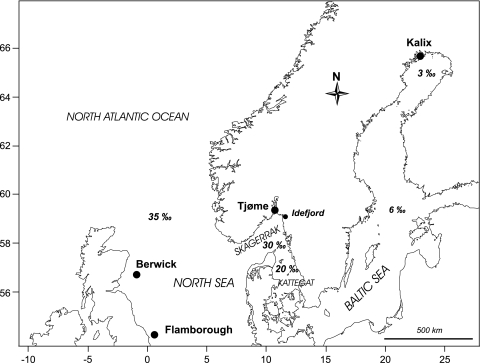
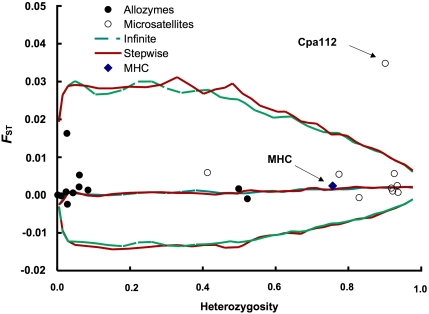
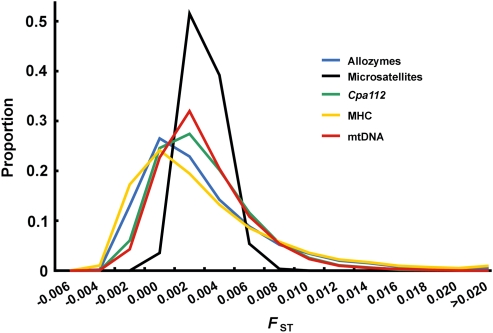
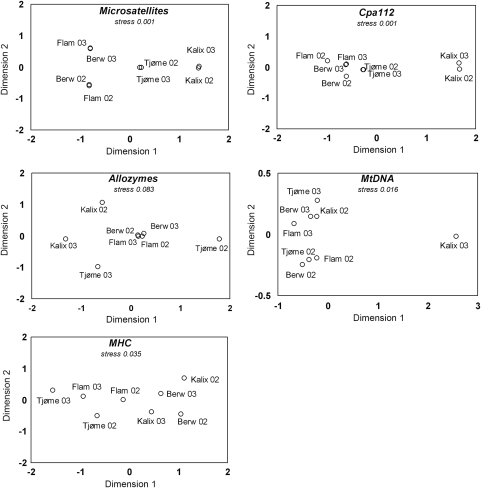
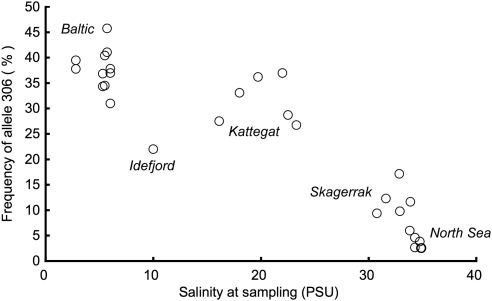
In many marine fish species, genetic population structure is typically weak because populations are large, evolutionarily young and have a high potential for gene flow. We tested whether genetic markers influenced by natural selection are more efficient than the presumed neutral genetic markers to detect population structure in Atlantic herring (Clupea harengus), a migratory pelagic species with large effective population sizes. We compared the spatial and temporal patterns of divergence and statistical power of three traditional genetic marker types, microsatellites, allozymes and mitochondrial DNA, with one microsatellite locus, Cpa112, previously shown to be influenced by divergent selection associated with salinity, and one locus located in the major histocompatibility complex class IIA (MHC-IIA) gene, using the same individuals across analyses. Samples were collected in 2002 and 2003 at two locations in the North Sea, one location in the Skagerrak and one location in the low-saline Baltic Sea. Levels of divergence for putatively neutral markers were generally low, with the exception of single outlier locus/sample combinations; microsatellites were the most statistically powerful markers under neutral expectations. We found no evidence of selection acting on the MHC locus. Cpa112, however, was highly divergent in the Baltic samples. Simulations addressing the statistical power for detecting population divergence showed that when using Cpa112 alone, compared with using eight presumed neutral microsatellite loci, sample sizes could be reduced by up to a tenth while still retaining high statistical power. Our results show that the loci influenced by selection can serve as powerful markers for detecting population structure in high gene-flow marine fish species.
Figures
References
-
- Allendorf FW, Seeb LW. Concordance of genetic divergence among sockeye salmon populations at allozyme, nuclear DNA, and mitochondrial DNA markers. Evolution. 2000;54:640–651. - PubMed
-
- Allendorf FW, Luikart G. Conservation and the Genetics of Populations. Blackwell Publishing: Malden, MA; 2007.
-
- Allendorf FW, England PR, Luikart G, Ritchie PA, Ryman N. Genetic effects of harvest on wild animal populations. Trends Ecol Evol. 2008;23:327–337. - PubMed
-
- Andersson L, Ryman N, Rosenberg R, Ståhl G. Genetic variability in Atlantic herring (Clupea harengus harengus): description of protein loci and population data. Hereditas. 1981;95:69–78.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
