

Evidence that CD8 T-cell homeostasis and function remain intact during murine pregnancy
- PMID: 20553337
- PMCID: PMC2996563
- DOI: 10.1111/j.1365-2567.2010.03316.x
Evidence that CD8 T-cell homeostasis and function remain intact during murine pregnancy
Abstract
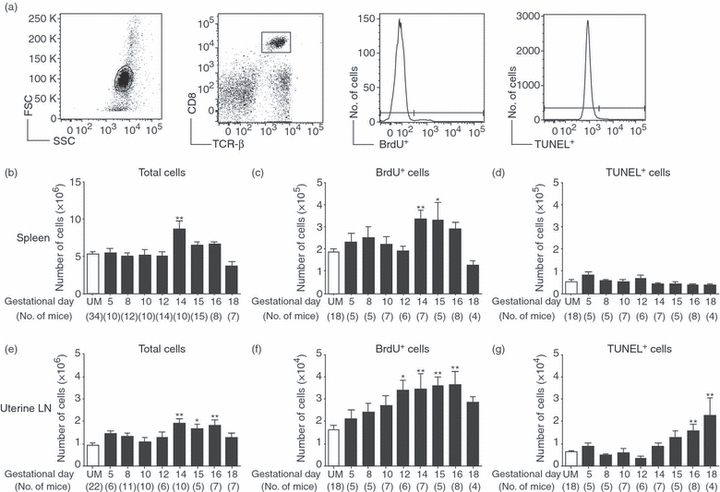
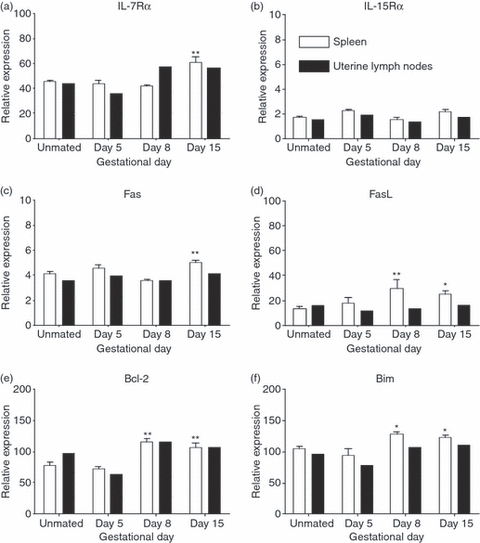
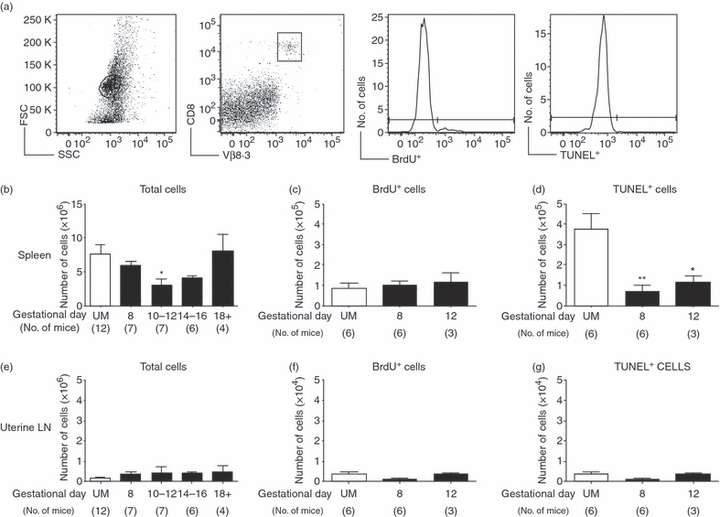
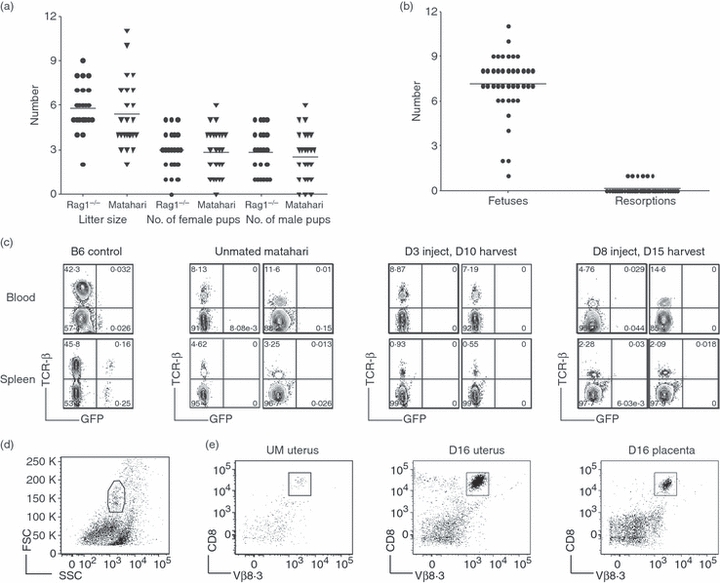
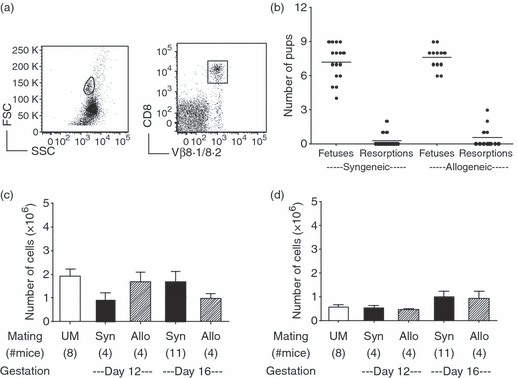
Evolving models of immune tolerance have challenged the view that the response of the maternal immune system to environmental or fetal antigens must be suppressed or deviated. CD8 T cells play a central role in the immune response to viruses and intracellular pathogens so the maintenance of both the number and function of these cells is critical to protect both the mother and fetus. We show that the numbers of maternal CD8 T cells in both the spleen and the uterine draining lymph nodes are transiently increased at mid-gestation and this correlates with enhanced CD8 T-cell proliferation and an increased relative expression of both pro-survival and pro-apoptotic molecules. In transgenic mice bearing T-cell antigen receptors specific for the male HY or allo-antigens, the transgenic CD8 T cells retain the ability to proliferate and function during pregnancy. Moreover, anti-HY T-cell receptor transgenic mice have normal numbers of male pups despite the presence of CD8 T cells at the maternal-fetal interface. These data suggest that pregnancy is a dynamic state in which CD8 T-cell turnover is increased while the function and ending size of the CD8 T-cell compartment are maintained.
© 2010 The Authors. Immunology © 2010 Blackwell Publishing Ltd.
Figures
Similar articles
-
Maternal PD-1 regulates accumulation of fetal antigen-specific CD8+ T cells in pregnancy.J Reprod Immunol. 2009 Jun;80(1-2):12-21. doi: 10.1016/j.jri.2008.12.001. Epub 2009 Apr 14. J Reprod Immunol. 2009. PMID: 19368976 Free PMC article.
-
Fetal expression of Fas ligand is necessary and sufficient for induction of CD8 T cell tolerance to the fetal antigen H-Y during pregnancy.J Immunol. 2005 Apr 15;174(8):4657-61. doi: 10.4049/jimmunol.174.8.4657. J Immunol. 2005. PMID: 15814689
-
Fas (CD95)-independent regulation of immune responses by antigen-specific CD4-CD8+ T cells.Int Immunol. 1996 May;8(5):675-81. doi: 10.1093/intimm/8.5.675. Int Immunol. 1996. PMID: 8671654
-
Utilising T cell receptor transgenic mice to define mechanisms of maternal T cell tolerance in pregnancy.J Reprod Immunol. 2010 Dec;87(1-2):1-13. doi: 10.1016/j.jri.2010.05.007. Epub 2010 Jul 8. J Reprod Immunol. 2010. PMID: 20615552 Review.
-
The New Old CD8+ T Cells in the Immune Paradox of Pregnancy.Front Immunol. 2021 Nov 16;12:765730. doi: 10.3389/fimmu.2021.765730. eCollection 2021. Front Immunol. 2021. PMID: 34868016 Free PMC article. Review.
Cited by
-
Decreased Expression of Cytotoxic Proteins in Decidual CD8+ T Cells in Preeclampsia.Biology (Basel). 2021 Oct 13;10(10):1037. doi: 10.3390/biology10101037. Biology (Basel). 2021. PMID: 34681139 Free PMC article.
-
Changes in T Cell and Dendritic Cell Phenotype from Mid to Late Pregnancy Are Indicative of a Shift from Immune Tolerance to Immune Activation.Front Immunol. 2017 Sep 15;8:1138. doi: 10.3389/fimmu.2017.01138. eCollection 2017. Front Immunol. 2017. PMID: 28966619 Free PMC article.
-
The landscape of decidual immune cells at the maternal-fetal interface in parturition and preterm birth.Inflamm Res. 2025 Mar 4;74(1):44. doi: 10.1007/s00011-025-02015-6. Inflamm Res. 2025. PMID: 40038160 Free PMC article. Review.
-
Synergies of Extracellular Vesicles and Microchimerism in Promoting Immunotolerance During Pregnancy.Front Immunol. 2022 Jul 1;13:837281. doi: 10.3389/fimmu.2022.837281. eCollection 2022. Front Immunol. 2022. PMID: 35844513 Free PMC article. Review.
-
Transient modification within a pool of CD4 T cells in the maternal spleen.Immunology. 2011 Nov;134(3):270-80. doi: 10.1111/j.1365-2567.2011.03486.x. Immunology. 2011. PMID: 21977997 Free PMC article.
References
-
- Baines M, Pross H, Millar K. Effects of pregnancy on the maternal lymphoid system in mice. Obstet Gynecol. 1977;50:457–61. - PubMed
-
- Hamilton M, Hellström I. Altered immune responses in pregnant mice. Transplantation. 1977;23:423–30. - PubMed
-
- Parmiani G, Invernizzi G. Depression of cellular immune response during syngeneic pregnancy as measured by the graft-versus-host reaction. Transplantation. 1975;19:363–8. - PubMed
-
- Jiang S, Vacchio M. Multiple mechanisms of peripheral T cell tolerance to the fetal “allograft”. J Immunol. 1998;160:3086–90. - PubMed
-
- Tafuri A, Alferink J, Möller P, Hämmerling G, Arnold B. T cell awareness of paternal alloantigens during pregnancy. Science. 1995;270:630–3. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
