

Single-molecule FRET of protein-nucleic acid and protein-protein complexes: surface passivation and immobilization
- PMID: 20554047
- PMCID: PMC3321382
- DOI: 10.1016/j.ymeth.2010.06.010
Single-molecule FRET of protein-nucleic acid and protein-protein complexes: surface passivation and immobilization
Abstract
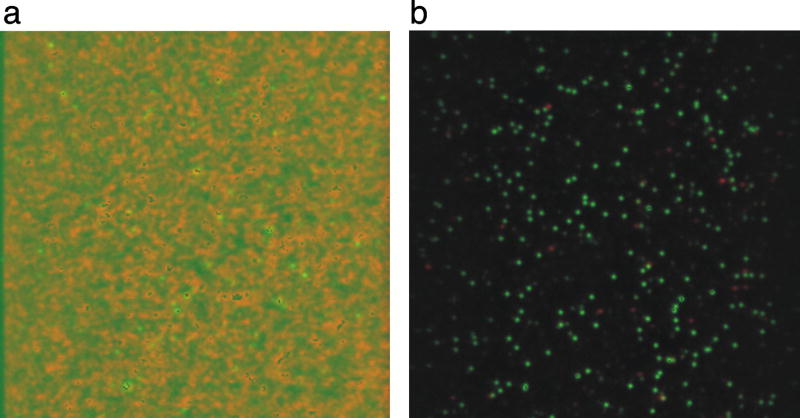
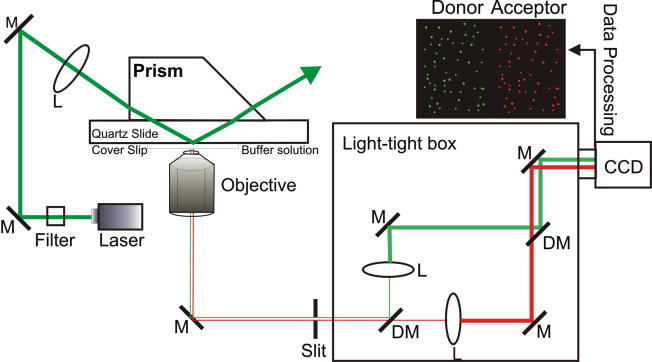
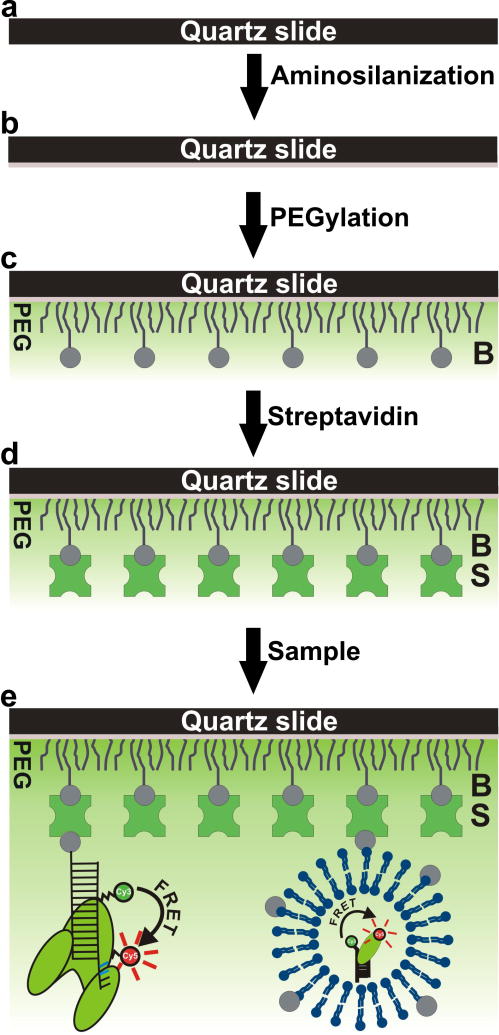
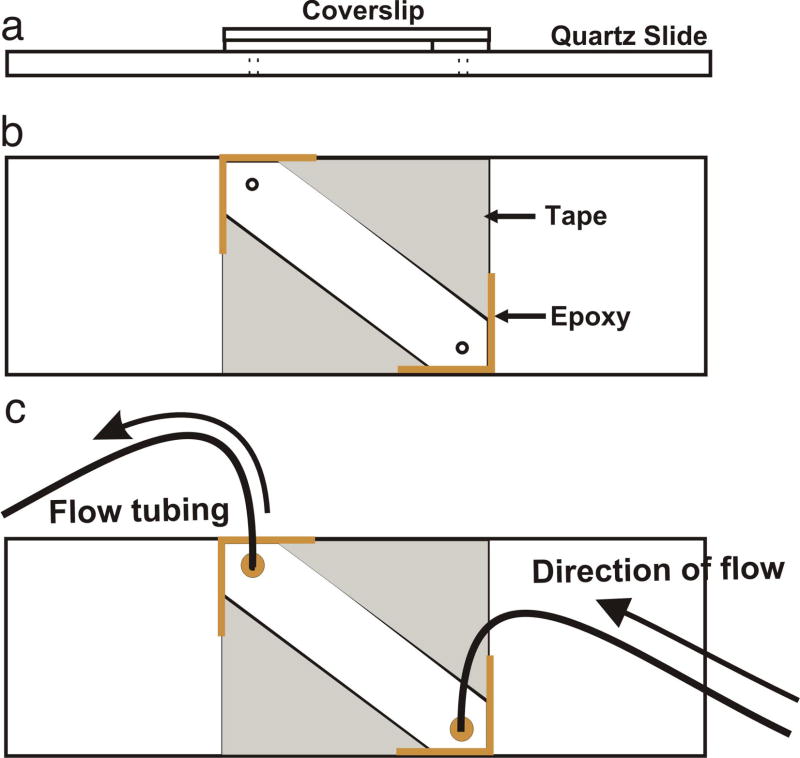
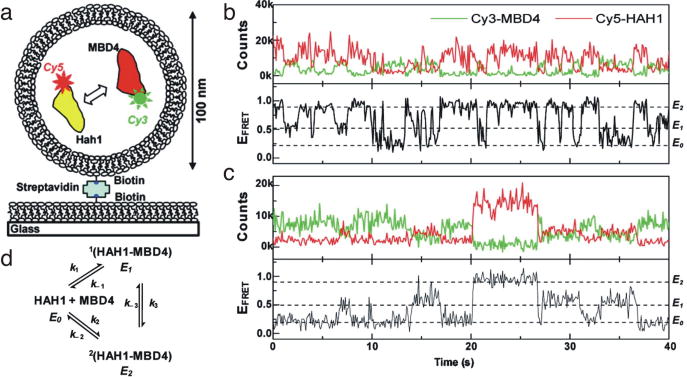
Single-molecule fluorescence spectroscopy reveals the real time dynamics that occur during biomolecular interactions that would otherwise be hidden by the ensemble average. It also removes the requirement to synchronize reactions, thus providing a very intuitive approach to study kinetics of biological systems. Surface immobilization is commonly used to increase observation times to the minute time scale, but it can be detrimental if the sample interacts non-specifically with the surface. Here, we review detailed protocols to prevent such interactions by passivating the surface or by trapping the molecules inside surface immobilized lipid vesicles. Finally, we discuss recent examples where these methods were applied to study the dynamics of important cellular processes at the single-molecule level.
Copyright © 2010 Elsevier Inc. All rights reserved.
Figures
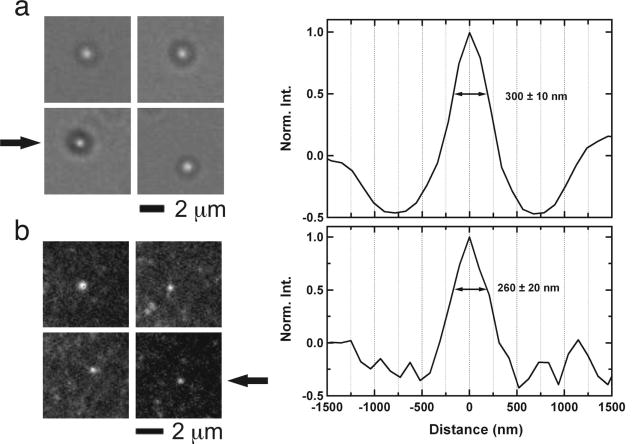
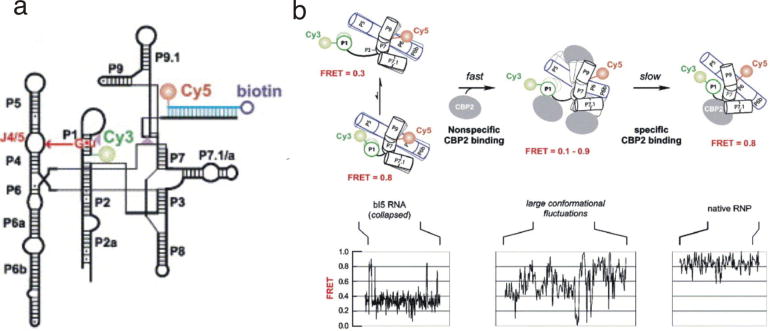
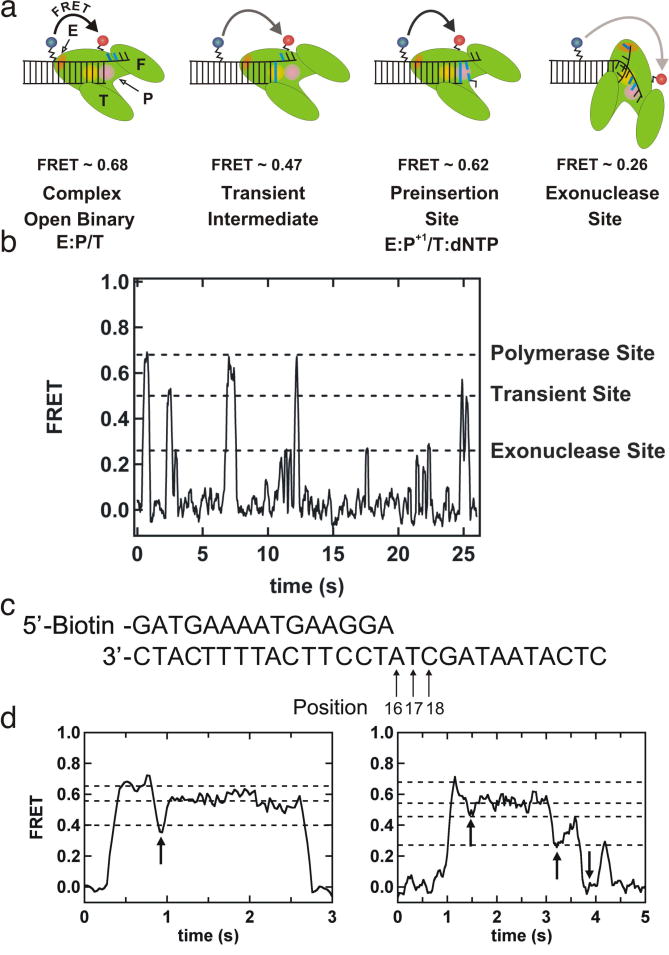
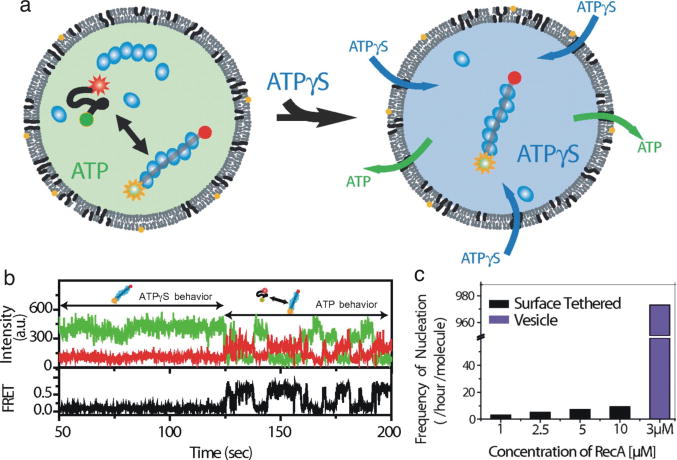
References
-
- Blanchard SC, Gonzalez RL, Kim HD, Chu S, Puglisi JD. Nat Struct Mol Biol. 2004;11:1008–14. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
