

Dynamic Network Connectivity: A new form of neuroplasticity
- PMID: 20554470
- PMCID: PMC2914830
- DOI: 10.1016/j.tics.2010.05.003
Dynamic Network Connectivity: A new form of neuroplasticity
Abstract
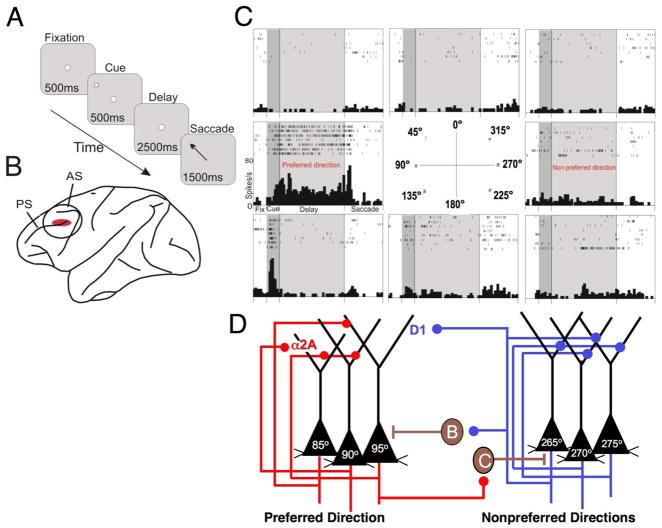
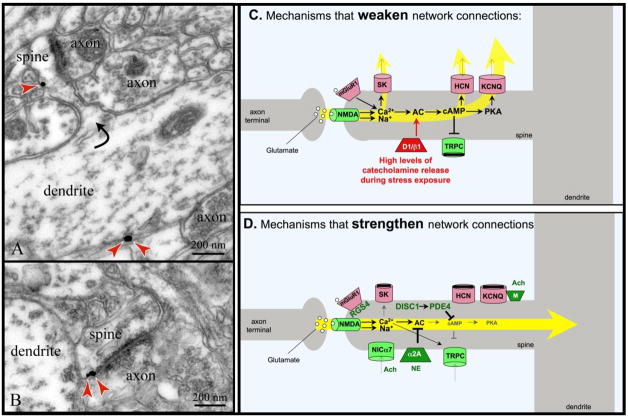
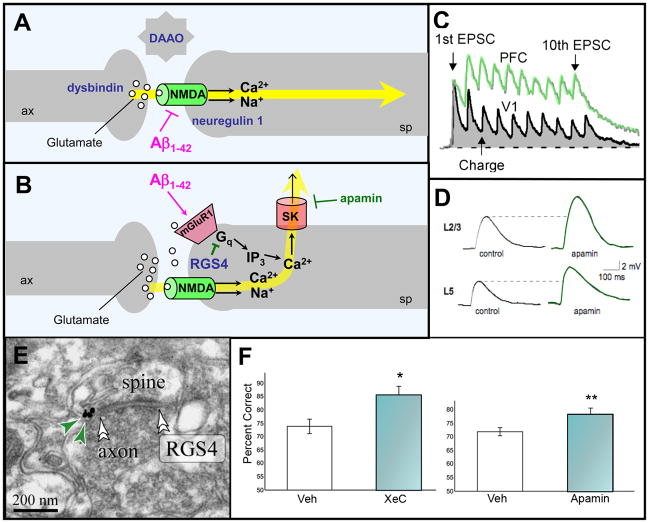
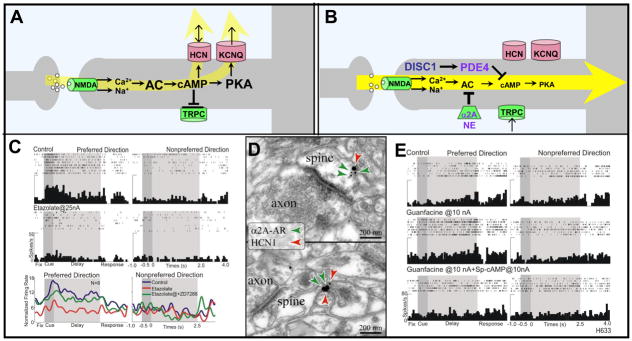
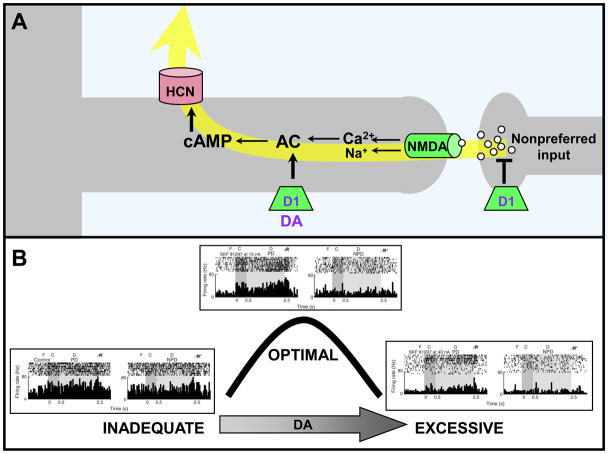
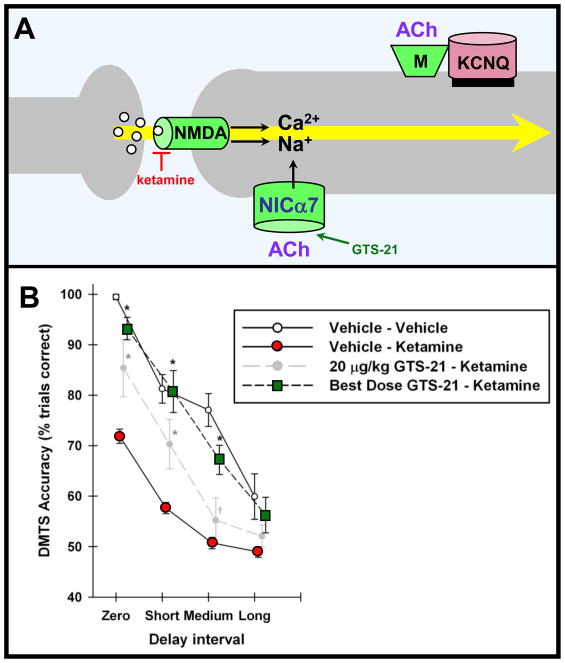
Prefrontal cortical (PFC) working memory functions depend on pyramidal cell networks that interconnect on dendritic spines. Recent research has revealed that the strength of PFC network connections can be rapidly and reversibly increased or decreased by molecular signaling events within slender, elongated spines: a process we term Dynamic Network Connectivity (DNC). This newly discovered form of neuroplasticity provides great flexibility in mental state, but also confers vulnerability and limits mental capacity. A remarkable number of genetic and/or environmental insults to DNC signaling cascades are associated with cognitive disorders such as schizophrenia and age-related cognitive decline. These insults can dysregulate network connections and erode higher cognitive abilities, leading to symptoms such as forgetfulness, susceptibility to interference, and disorganized thought and behavior.
Copyright 2010 Elsevier Ltd. All rights reserved.
Figures
References
-
- Moore TL, et al. Executive system dysfunction occurs as early as middle-age in the rhesus monkey. Neurobiol Aging. 2006;27:1484–1493. - PubMed
-
- Weinberger DR, et al. Physiologic dysfunction of dorsolateral prefrontal cortex in schizophrenia. I. Regional cerebral blood flow evidence. Archives General Psychiatry. 1986;43:114–124. - PubMed
-
- Rubia K, et al. Hypofrontality in Attention Deficit Hyperactivity Disorder during higher-order motor control: A study with functional MRI. Am J Psychiatry. 1999;156:891–896. - PubMed
-
- Blumberg HP, et al. A functional magnetic resonance imaging study of bipolar disorder: state- and trait-related dysfunction in ventral prefrontal cortices. Arch Gen Psychiatry. 2003;60:601–609. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
