

Functional properties of the human cytomegalovirus IE86 protein required for transcriptional regulation and virus replication
- PMID: 20554773
- PMCID: PMC2919014
- DOI: 10.1128/JVI.00327-10
Functional properties of the human cytomegalovirus IE86 protein required for transcriptional regulation and virus replication
Abstract
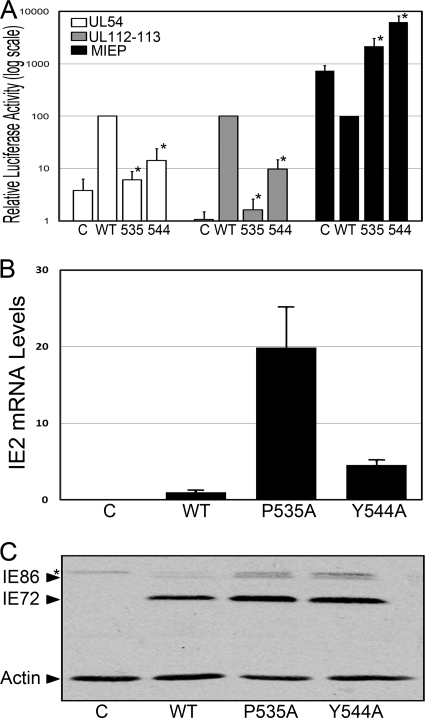
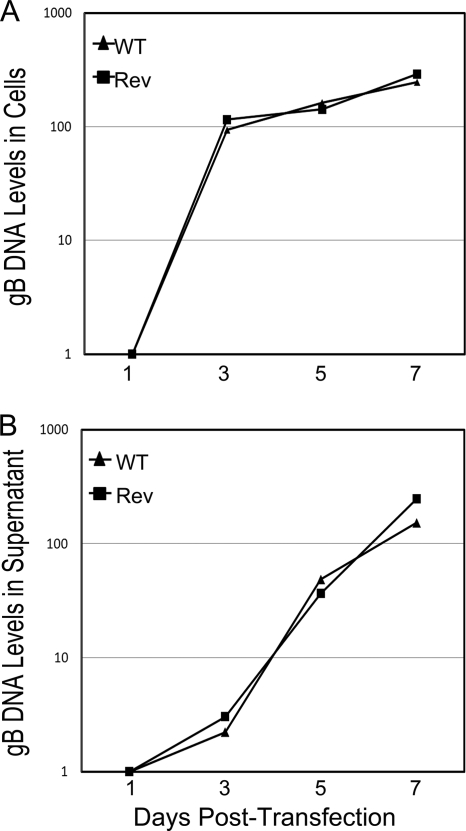
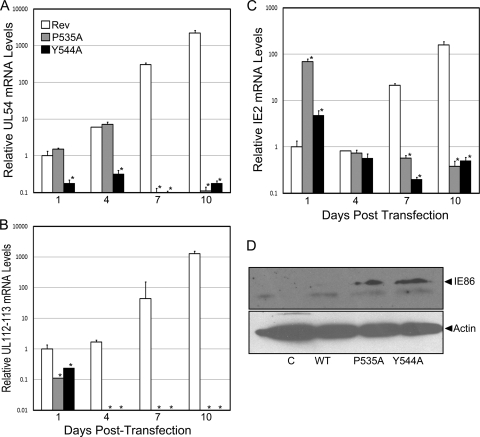
The human cytomegalovirus (HCMV) IE86 protein is essential for HCMV replication due to its ability to transactivate critical viral early promoters. In the current study, we performed a comprehensive mutational analysis between amino acids (aa) 535 and 545 of IE86 and assessed the impact of these mutations on IE86-mediated transcriptional activation. Using transient assays and complementing analysis with recombinant HCMV clones, we show that single amino acid mutations differentially impair the ability of IE86 to mediate transactivation of essential early gene promoters. The conserved tyrosine at amino acid 544 is critical for activation of the UL54 promoter in vitro and in the context of the viral genome. In contrast, mutation of the proline at position 535 disrupted activation of the UL54 promoter in transient assays but displayed activity similar to that of wild-type (WT) IE86 when assessed in the genomic context. To examine the underlying mechanism of this differential effect, glutathione S-transferase (GST) pulldown assays were performed, revealing that Y544 is critical for binding to the TATA binding protein (TBP), suggesting that this interaction is likely necessary for the ability of IE86 to activate the UL54 promoter. In contrast, mutation of either P535 or Y544 disrupted activation of the UL112-113 promoter both in vitro and in vivo, suggesting that interaction with TBP is not sufficient for IE86-mediated activation of this early promoter. Together, these studies demonstrate that IE86 activates early promoters by distinct mechanisms.
Figures



References
-
- Ahn, J. H., C. J. Chiou, and G. S. Hayward. 1998. Evaluation and mapping of the DNA binding and oligomerization domains of the IE2 regulatory protein of human cytomegalovirus using yeast one and two hybrid interaction assays. Gene 210:25-36. - PubMed
-
- Asmar, J., L. Wiebusch, M. Truss, and C. Hagemeier. 2004. The putative zinc finger of the human cytomegalovirus IE2 86-kilodalton protein is dispensable for DNA binding and autorepression, thereby demarcating a concise core domain in the C terminus of the protein. J. Virol. 78:11853-11864. - PMC - PubMed
-
- Barrasa, M. I., N. Harel, Y. Yu, and J. C. Alwine. 2003. Strain variations in single amino acids of the 86-kilodalton human cytomegalovirus major immediate-early protein (IE2) affect its functional and biochemical properties: implications of dynamic protein conformation. J. Virol. 77:4760-4772. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
