

A pathologic cascade leading to synaptic dysfunction in alpha-synuclein-induced neurodegeneration
- PMID: 20554859
- PMCID: PMC2901533
- DOI: 10.1523/JNEUROSCI.1091-10.2010
A pathologic cascade leading to synaptic dysfunction in alpha-synuclein-induced neurodegeneration
Abstract
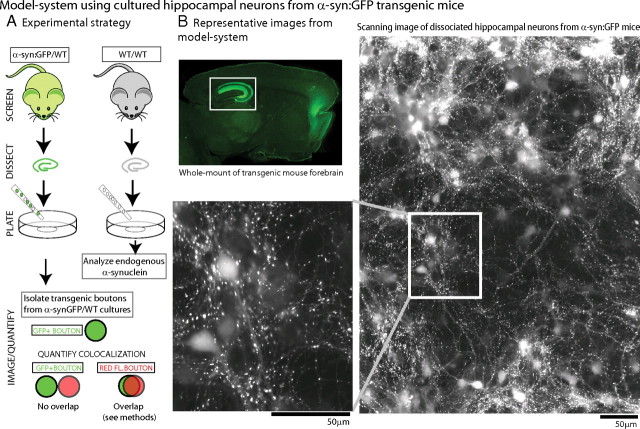
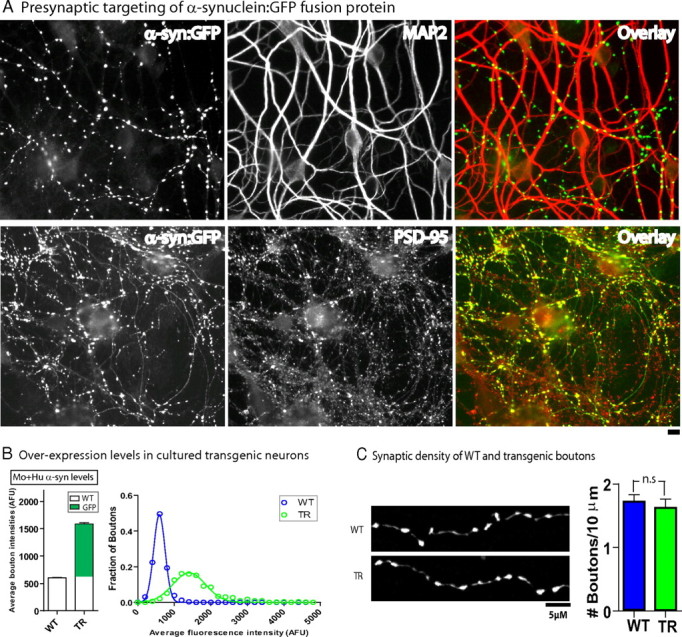
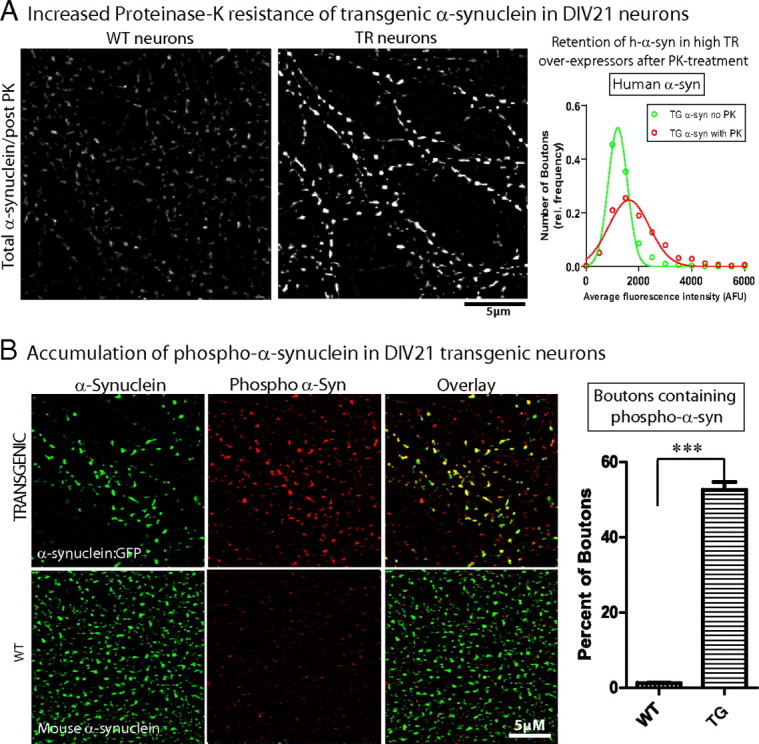
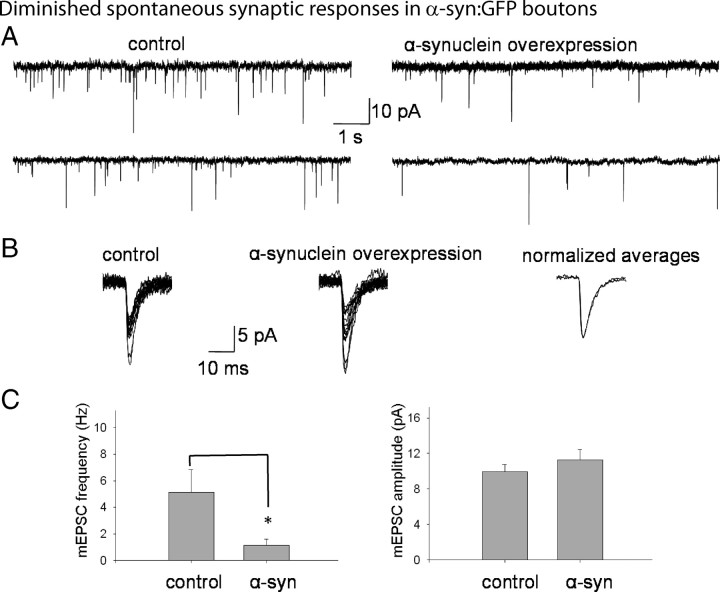
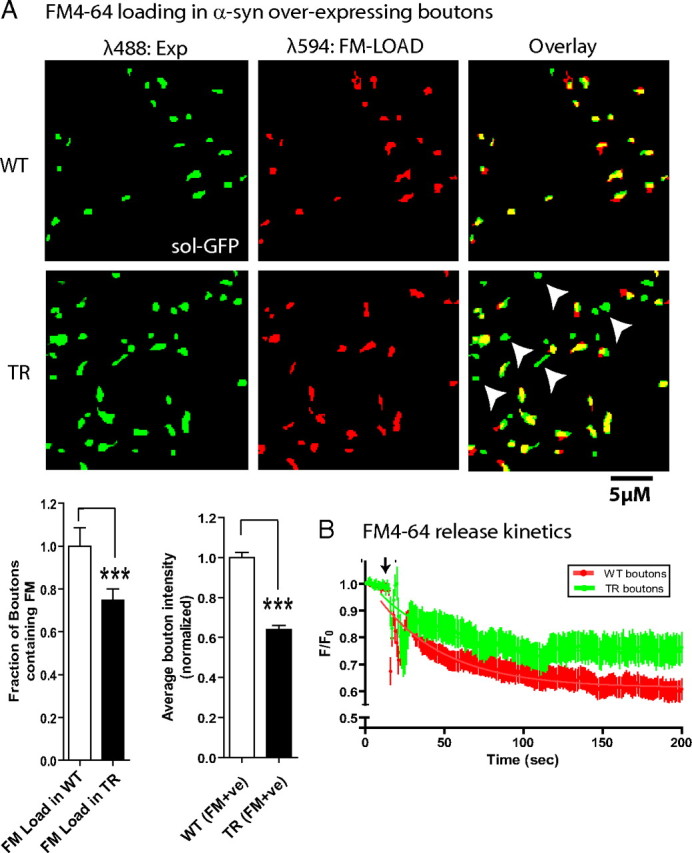
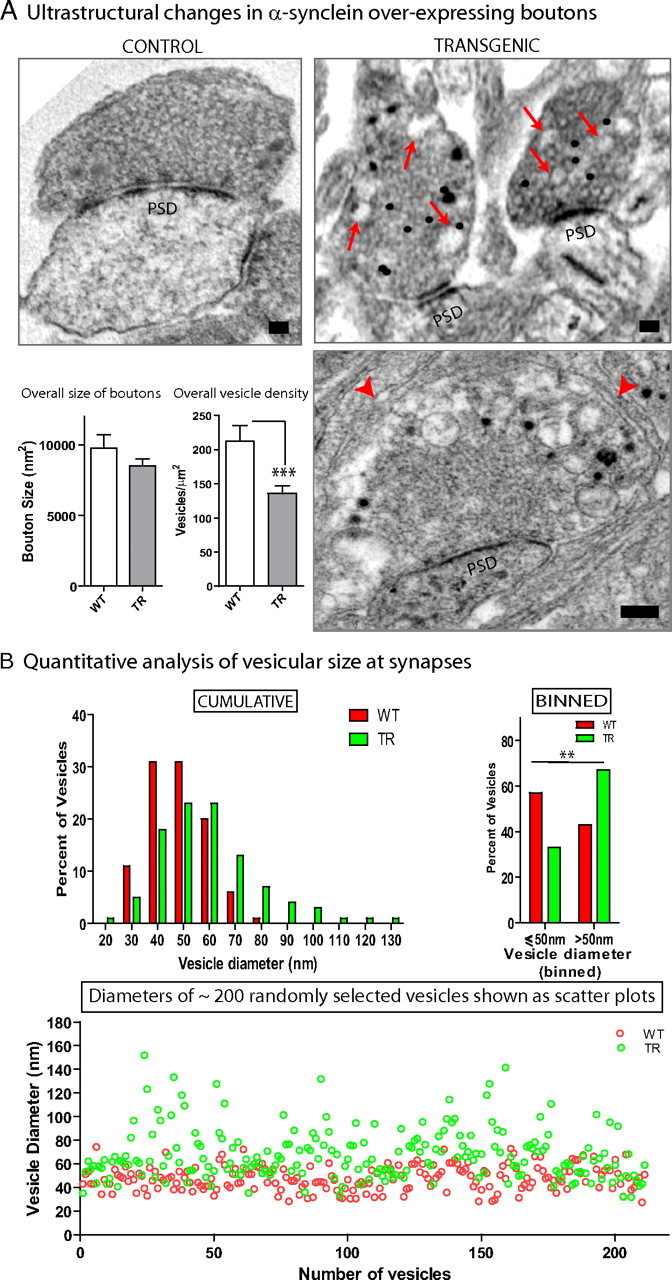
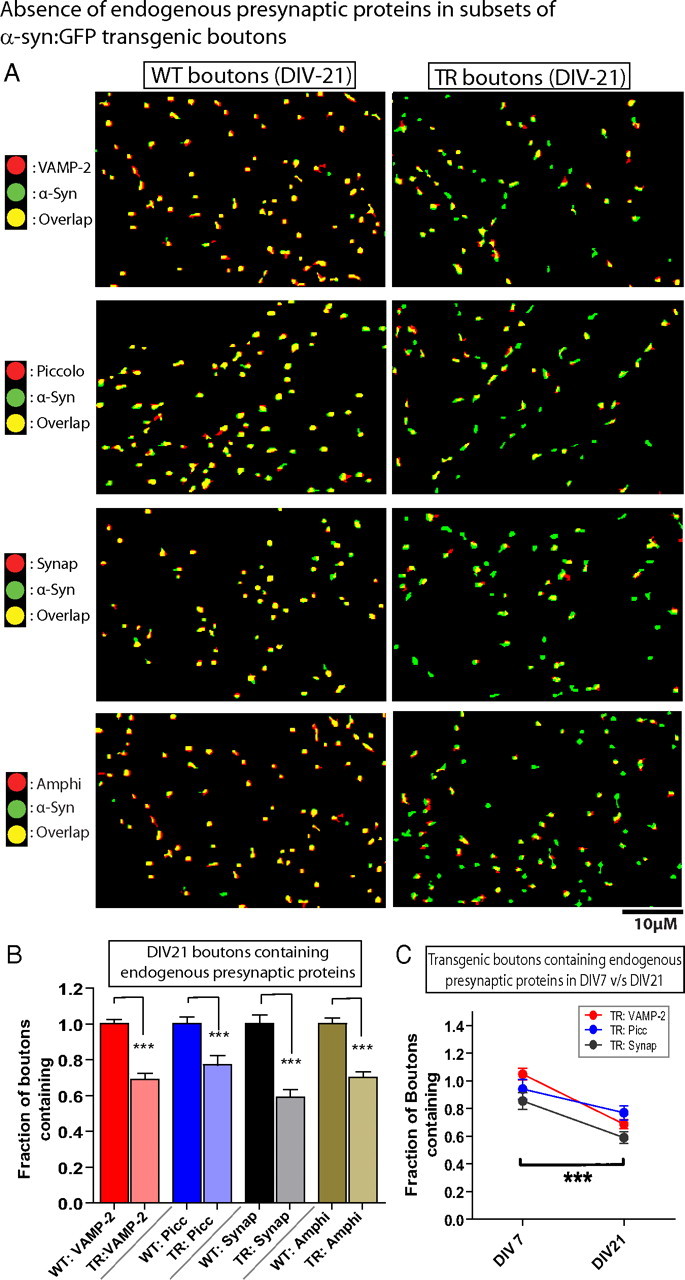
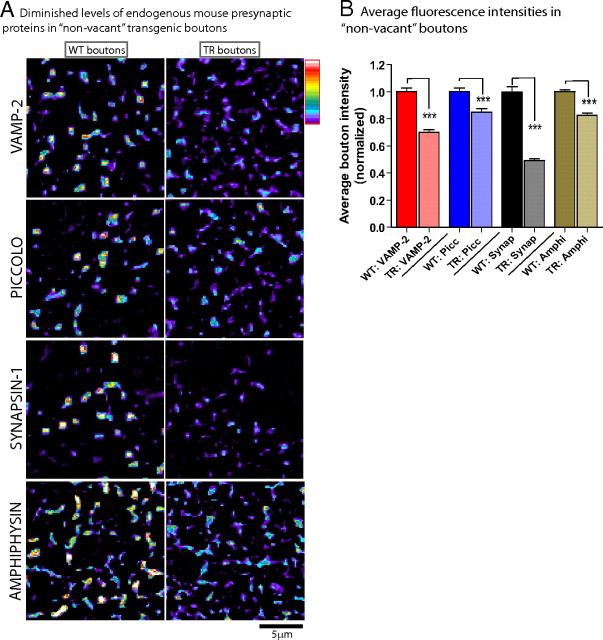
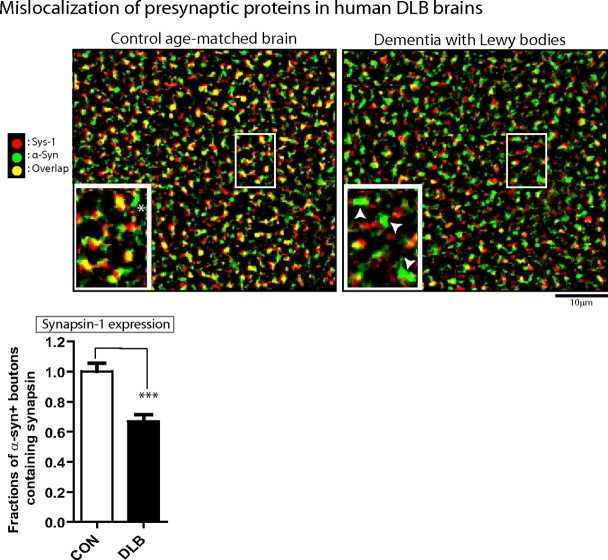
Several neurodegenerative diseases are typified by intraneuronal alpha-synuclein deposits, synaptic dysfunction, and dementia. While even modest alpha-synuclein elevations can be pathologic, the precise cascade of events induced by excessive alpha-synuclein and eventually culminating in synaptotoxicity is unclear. To elucidate this, we developed a quantitative model system to evaluate evolving alpha-synuclein-induced pathologic events with high spatial and temporal resolution, using cultured neurons from brains of transgenic mice overexpressing fluorescent-human-alpha-synuclein. Transgenic alpha-synuclein was pathologically altered over time and overexpressing neurons showed striking neurotransmitter release deficits and enlarged synaptic vesicles; a phenotype reminiscent of previous animal models lacking critical presynaptic proteins. Indeed, several endogenous presynaptic proteins involved in exocytosis and endocytosis were undetectable in a subset of transgenic boutons ("vacant synapses") with diminished levels in the remainder, suggesting that such diminutions were triggering the overall synaptic pathology. Similar synaptic protein alterations were also retrospectively seen in human pathologic brains, highlighting potential relevance to human disease. Collectively the data suggest a previously unknown cascade of events where pathologic alpha-synuclein leads to a loss of a number of critical presynaptic proteins, thereby inducing functional synaptic deficits.
Figures
References
-
- Aarsland D, Beyer MK, Kurz MW. Dementia in Parkinson's disease. Curr Opin Neurol. 2008;21:676–682. - PubMed
-
- Abeliovich A, Schmitz Y, Fariñas I, Choi-Lundberg D, Ho WH, Castillo PE, Shinsky N, Verdugo JM, Armanini M, Ryan A, Hynes M, Phillips H, Sulzer D, Rosenthal A. Mice lacking alpha-synuclein display functional deficits in the nigrostriatal dopamine system. Neuron. 2000;25:239–252. - PubMed
-
- Anderson JP, Walker DE, Goldstein JM, de Laat R, Banducci K, Caccavello RJ, Barbour R, Huang J, Kling K, Lee M, Diep L, Keim PS, Shen X, Chataway T, Schlossmacher MG, Seubert P, Schenk D, Sinha S, Gai WP, Chilcote TJ. Phosphorylation of Ser-129 is the dominant pathological modification of alpha-synuclein in familial and sporadic Lewy body disease. J Biol Chem. 2006;281:29739–29752. - PubMed
-
- Chandra S, Gallardo G, Fernández-Chacón R, Schlüter OM, Südhof TC. Alpha-synuclein cooperates with CSPalpha in preventing neurodegeneration. Cell. 2005;123:383–396. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases