

CaMKII "autonomy" is required for initiating but not for maintaining neuronal long-term information storage
- PMID: 20554872
- PMCID: PMC2891520
- DOI: 10.1523/JNEUROSCI.1469-10.2010
CaMKII "autonomy" is required for initiating but not for maintaining neuronal long-term information storage
Abstract
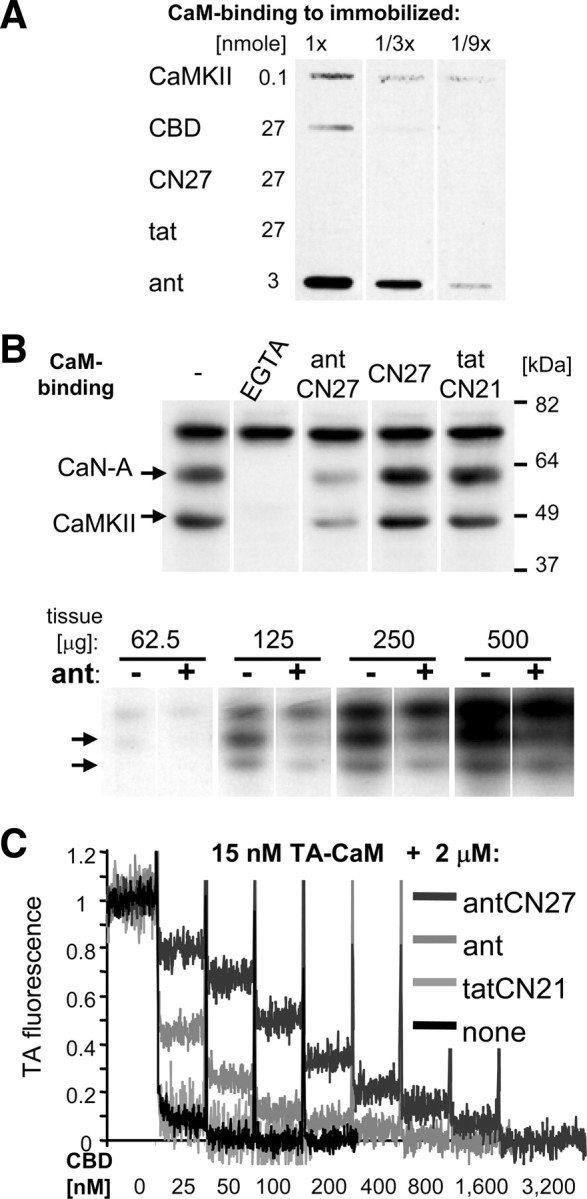
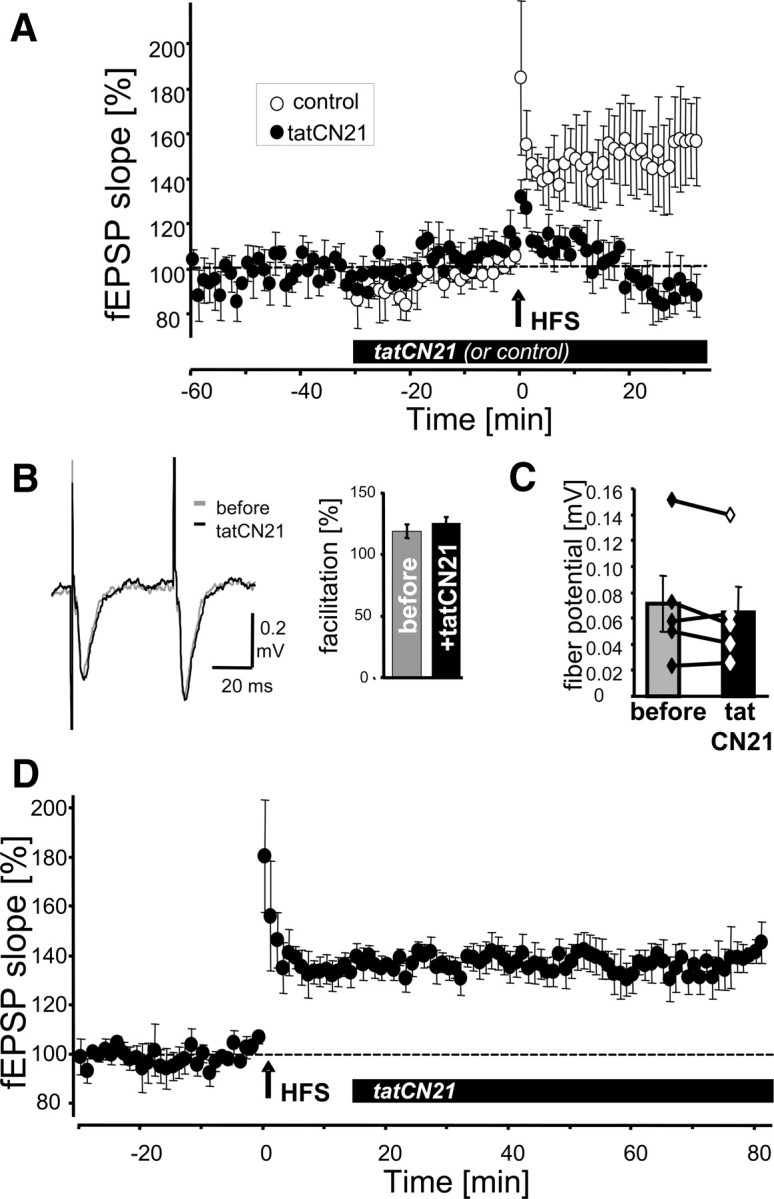
Ca(2+)/calmodulin (CaM)-dependent protein kinase II (CaMKII) "autonomy" (T286-autophosphorylation-induced Ca(2+)-independent activity) is required for long-term potentiation (LTP) and for learning and memory, as demonstrated by CaMKII T286A mutant mice. The >20-year-old hypothesis that CaMKII stimulation is required for LTP induction, while CaMKII autonomy is required for LTP maintenance was recently supported using the cell-penetrating fusion-peptide inhibitor antCN27. However, we demonstrate here that ant/penetratin fusion to CN27 compromised CaMKII-selectivity, by enhancing a previously unnoticed direct binding of CaM to ant/penetratin. In contrast to antCN27, the improved cell-penetrating inhibitor tatCN21 (5 mum) showed neither CaM binding nor inhibition of basal synaptic transmission. In vitro, tatCN21 inhibited stimulated and autonomous CaMKII activity with equal potency. In rat hippocampal slices, tatCN21 inhibited LTP induction, but not LTP maintenance. Correspondingly, tatCN21 also inhibited learning, but not memory storage or retrieval in a mouse in vivo model. Thus, CaMKII autonomy provides a short-term molecular memory that is important in the signal computation leading to memory formation, but is not required as long-term memory store.
Figures



References
-
- Aarts M, Liu Y, Liu L, Besshoh S, Arundine M, Gurd JW, Wang YT, Salter MW, Tymianski M. Treatment of ischemic brain damage by perturbing NMDA receptor-PSD-95 protein interactions. Science. 2002;298:846–850. - PubMed
-
- Barria A, Malinow R. NMDA receptor subunit composition controls synaptic plasticity by regulating binding to CaMKII. Neuron. 2005;48:289–301. - PubMed
-
- Bayer KU, De Koninck P, Leonard AS, Hell JW, Schulman H. Interaction with the NMDA receptor locks CaMKII in an active conformation. Nature. 2001;411:801–805. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous