

Structural conservation predominates over sequence variability in the crown of HIV type 1's V3 loop
- PMID: 20560796
- PMCID: PMC2932551
- DOI: 10.1089/aid.2009.0254
Structural conservation predominates over sequence variability in the crown of HIV type 1's V3 loop
Abstract
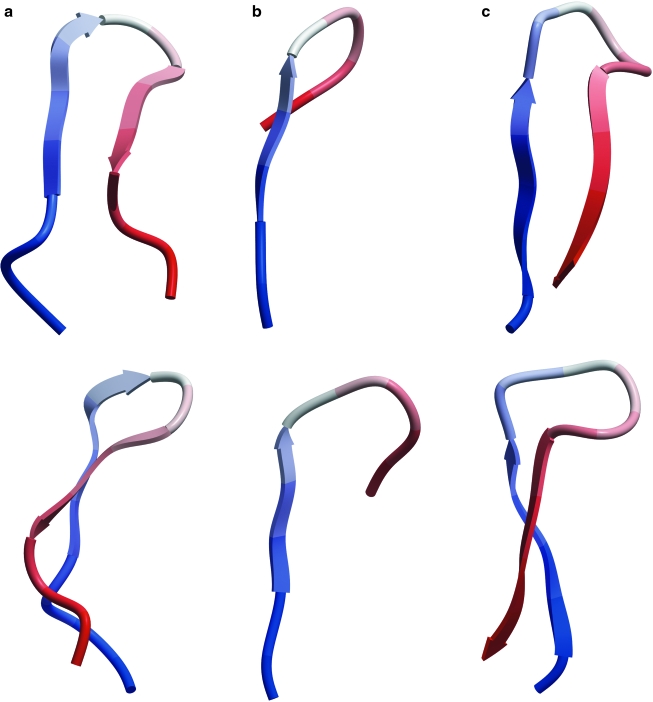
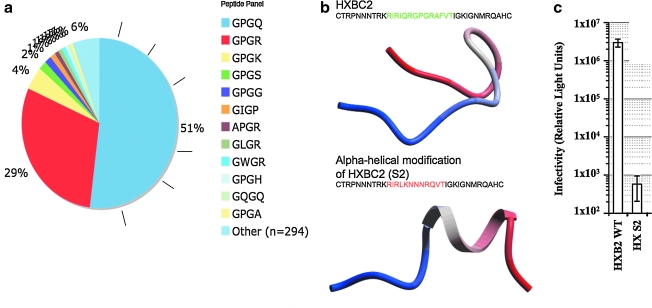
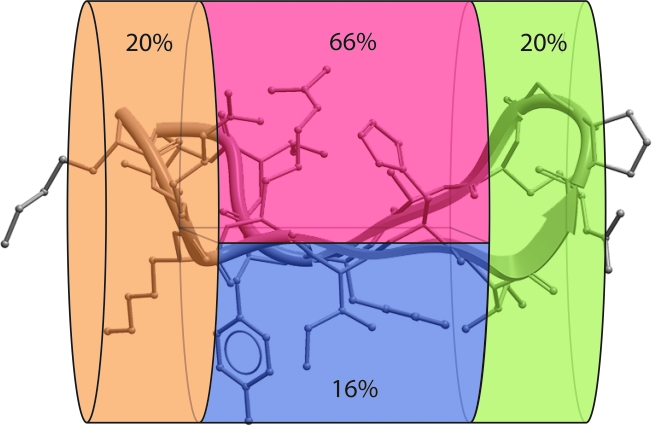
The diversity of HIV-1 is a confounding problem for vaccine design, as the human immune response appears to favor poor or strain-specific responses to any given HIV-1 virus strain. A significant portion of this diversity is manifested as sequence variability in the loops of HIV-1's surface envelope glycoprotein. Here we show that the most variable sequence positions in the third variable (V3) loop crown cluster to a small zone on the surface of one face of the V3 loop ss-hairpin conformation. These results provide a novel visualization of the gp120 V3 loop, specifically demonstrating a surprising preponderance of conserved three-dimensional structure in a highly sequence-variable region. From a structural point of view, there appears to be less diversity in this region of the HIV-1 "principle neutralizing domain" than previously appreciated.
Figures
References
-
- Trkola A. Dragic T. Arthos J, et al. CD4-dependent, antibody-sensitive interactions between HIV-1 and its co-receptor CCR-5. Nature. 1996;384(6605):184–187. - PubMed
-
- Hill C. Deng H. Unutmaz D, et al. Envelope glycoproteins from human immunodeficiency virus types 1 and 2 and simian immunodeficiency virus can use human CCR5 as a coreceptor for viral entry and make direct CD4-dependent interactions with this chemokine receptor. J Virol. 1997;71(9):6296–6304. - PMC - PubMed
-
- Leitner T, editor; Foley B, editor; Hahn B, et al., editors. HIV Sequence Compendium. Los Alamos National Laboratory, NM: Theoretical Biology and Biophysics Group; 2005. No. LA-UR 06-0680.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
