

Kinetic evidence for the presence of putative germination receptors in Clostridium difficile spores
- PMID: 20562307
- PMCID: PMC2916422
- DOI: 10.1128/JB.00488-10
Kinetic evidence for the presence of putative germination receptors in Clostridium difficile spores
Abstract
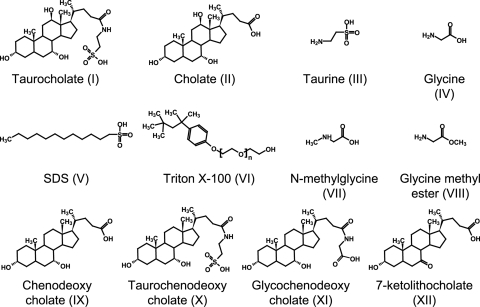
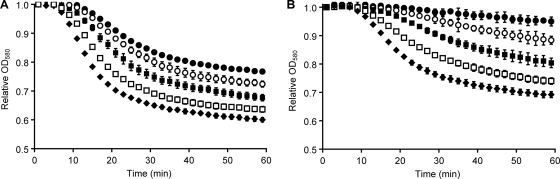
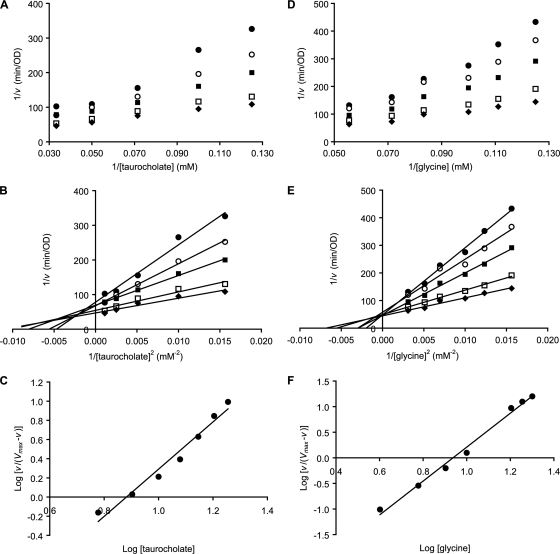
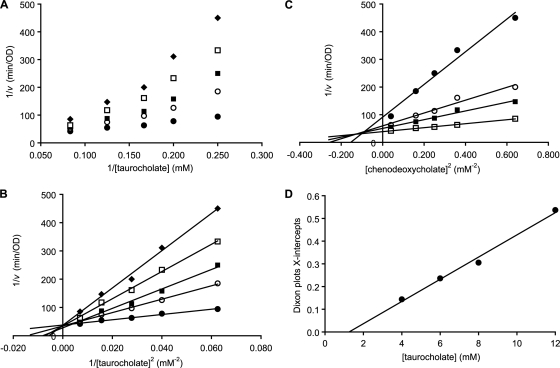
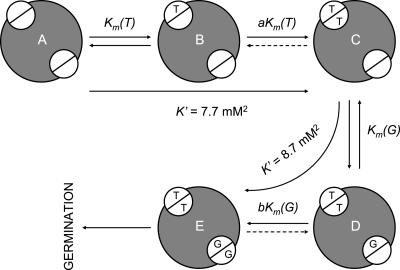
Clostridium difficile is a spore-forming bacterium that causes Clostridium difficile-associated disease (CDAD). Intestinal microflora keeps C. difficile in the spore state and prevents colonization. Following antimicrobial treatment, the microflora is disrupted, and C. difficile spores germinate in the intestines. The resulting vegetative cells are believed to fill empty niches left by the depleted microbial community and establish infection. Thus, germination of C. difficile spores is the first required step in CDAD. Interestingly, C. difficile genes encode most known spore-specific protein necessary for germination, except for germination (Ger) receptors. Even though C. difficile Ger receptors have not been identified, taurocholate (a bile salt) and glycine (an amino acid) have been shown to be required for spore germination. Furthermore, chenodeoxycholate, another bile salt, can inhibit taurocholate-induced C. difficile spore germination. In the present study, we examined C. difficile spore germination kinetics to determine whether taurocholate acts as a specific germinant that activates unknown germination receptors or acts nonspecifically by disrupting spores' membranes. Kinetic analysis of C. difficile spore germination suggested the presence of distinct receptors for taurocholate and glycine. Furthermore, taurocholate, glycine, and chenodeoxycholate seem to bind to C. difficile spores through a complex mechanism, where both receptor homo- and heterocomplexes are formed. The kinetic data also point to an ordered sequential progression of binding where taurocholate must be recognized first before detection of glycine can take place. Finally, comparing calculated kinetic parameters with intestinal concentrations of the two germinants suggests a mechanism for the preferential germination of C. difficile spores in antibiotic-treated individuals.
Figures
Similar articles
-
Inhibiting the initiation of Clostridium difficile spore germination using analogs of chenodeoxycholic acid, a bile acid.J Bacteriol. 2010 Oct;192(19):4983-90. doi: 10.1128/JB.00610-10. Epub 2010 Jul 30. J Bacteriol. 2010. PMID: 20675492 Free PMC article.
-
Mapping interactions between germinants and Clostridium difficile spores.J Bacteriol. 2011 Jan;193(1):274-82. doi: 10.1128/JB.00980-10. Epub 2010 Oct 22. J Bacteriol. 2011. PMID: 20971909 Free PMC article.
-
Hierarchical recognition of amino acid co-germinants during Clostridioides difficile spore germination.Anaerobe. 2018 Feb;49:41-47. doi: 10.1016/j.anaerobe.2017.12.001. Epub 2017 Dec 6. Anaerobe. 2018. PMID: 29221987 Free PMC article.
-
A Revised Understanding of Clostridioides difficile Spore Germination.Trends Microbiol. 2020 Sep;28(9):744-752. doi: 10.1016/j.tim.2020.03.004. Epub 2020 Apr 23. Trends Microbiol. 2020. PMID: 32781028 Review.
-
Clostridium difficile spore biology: sporulation, germination, and spore structural proteins.Trends Microbiol. 2014 Jul;22(7):406-16. doi: 10.1016/j.tim.2014.04.003. Epub 2014 May 7. Trends Microbiol. 2014. PMID: 24814671 Free PMC article. Review.
Cited by
-
Effect of the Synthetic Bile Salt Analog CamSA on the Hamster Model of Clostridium difficile Infection.Antimicrob Agents Chemother. 2018 Sep 24;62(10):e02251-17. doi: 10.1128/AAC.02251-17. Print 2018 Oct. Antimicrob Agents Chemother. 2018. PMID: 30012758 Free PMC article.
-
The Design, Synthesis, and Characterizations of Spore Germination Inhibitors Effective against an Epidemic Strain of Clostridium difficile.J Med Chem. 2018 Aug 9;61(15):6759-6778. doi: 10.1021/acs.jmedchem.8b00632. Epub 2018 Jul 30. J Med Chem. 2018. PMID: 30004695 Free PMC article.
-
The design, synthesis, and inhibition of Clostridioides difficile spore germination by acyclic and bicyclic tertiary amide analogs of cholate.Eur J Med Chem. 2023 Dec 5;261:115788. doi: 10.1016/j.ejmech.2023.115788. Epub 2023 Sep 4. Eur J Med Chem. 2023. PMID: 37703709 Free PMC article.
-
The Impact of YabG Mutations on Clostridioides difficile Spore Germination and Processing of Spore Substrates.Mol Microbiol. 2024 Oct;122(4):534-548. doi: 10.1111/mmi.15316. Epub 2024 Sep 11. Mol Microbiol. 2024. PMID: 39258427
-
Inhibiting the initiation of Clostridium difficile spore germination using analogs of chenodeoxycholic acid, a bile acid.J Bacteriol. 2010 Oct;192(19):4983-90. doi: 10.1128/JB.00610-10. Epub 2010 Jul 30. J Bacteriol. 2010. PMID: 20675492 Free PMC article.
References
-
- Abel-Santos, E., and T. Dodatko. 2007. Differential nucleoside recognition during Bacillus cereus 569 (ATCC 10876) spore germination. New J. Chem. 31:748-755.
-
- Ahlman, B., C.-E. Leijonmarck, C. Lind, E. Vinnars, and J. Wernerman. 1993. Free amino acids in biopsy specimens from the human colonic mucosa. J. Surgic. Res. 55:647-653. - PubMed
-
- Akoachere, M., R. C. Squires, A. M. Nour, L. Angelov, J. Brojatsch, and E. V. Abel-Santos. 2007. Identification of an in vivo inhibitor of Bacillus anthracis Sterne spore germination. J. Biol. Chem. 282:12112-12118. - PubMed
-
- Alberto, F., V. Broussolle, D. R. Mason, F. Carlin, and M. W. Peck. 2003. Variability in spore germination response by strains of proteolytic Clostridium botulinum types A, B and F. Lett. Appl. Microbiol. 36:41-45. - PubMed
-
- Arslan, H., E. K. Inci, O. K. Azap, H. Karakayali, A. Torgay, and M. Haberal. 2007. Etiologic agents of diarrhea in solid organ recipients. Transpl. Infect. Dis. 9:270-275. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
