

P2X4 receptors mediate PGE2 release by tissue-resident macrophages and initiate inflammatory pain
- PMID: 20562826
- PMCID: PMC2910276
- DOI: 10.1038/emboj.2010.126
P2X4 receptors mediate PGE2 release by tissue-resident macrophages and initiate inflammatory pain
Abstract
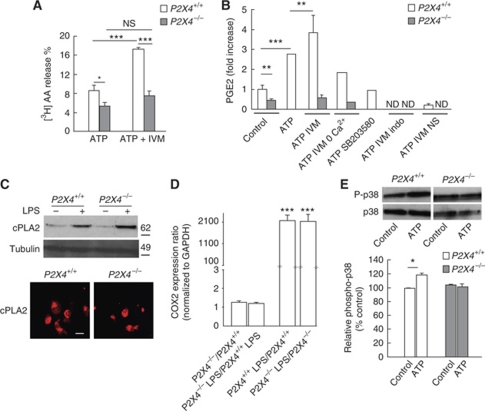
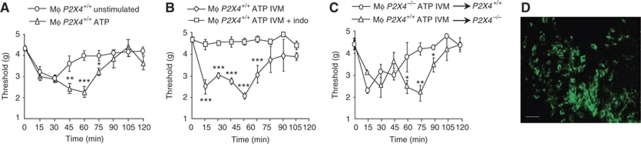
Prostaglandin E2 (PGE2) is a key mediator of inflammation and contributes to pain hypersensitivity by promoting sensory neurons hyperexcitability. PGE2 synthesis results from activation of a multi-step enzymatic cascade that includes cyclooxygenases (COXs), the main targets of non-steroidal anti-inflammatory drugs (NSAIDs). Although NSAIDs are widely prescribed to reduce inflammatory symptoms such as swelling and pain, associated harmful side effects restrict their long-term use. Therefore, finding new drugs that limit PG production represents an important therapeutic issue. In response to peripheral inflammatory challenges, mice lacking the ATP-gated P2X4 channel (P2X4R) do not develop pain hypersensitivity and show a complete absence of inflammatory PGE2 in tissue exudates. In resting conditions, tissue-resident macrophages constitutively express P2X4R. Stimulating P2X4R in macrophages triggers calcium influx and p38 MAPK phosphorylation, resulting in cytosolic PLA2 (cPLA2) activation and COX-dependent release of PGE2. In naive animals, pain hypersensitivity was elicited by transfer into the paw of ATP-primed macrophages from wild type, but not P2X4R-deficient mice. Thus, P2X4Rs are specifically involved in inflammatory-mediated PGE2 production and might therefore represent useful therapeutic targets.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures




Similar articles
-
Exogenous ATP modulates PGE2 release in macrophages through sustained phosphorylation of CDK9 and p38 MAPK.J Leukoc Biol. 2021 Oct;110(4):663-677. doi: 10.1002/JLB.3A1219-697RR. Epub 2021 Jan 13. J Leukoc Biol. 2021. PMID: 33438260
-
Role of mitogen-activated protein kinases in influenza virus induction of prostaglandin E2 from arachidonic acid in bronchial epithelial cells.Clin Exp Allergy. 2003 Sep;33(9):1244-51. doi: 10.1046/j.1365-2222.2003.01750.x. Clin Exp Allergy. 2003. PMID: 12956746
-
Lung inflammation caused by adenosine-5'-triphosphate is mediated via Ca2+/PKCs-dependent COX-2/PGE2 induction.Int J Biochem Cell Biol. 2013 Aug;45(8):1657-68. doi: 10.1016/j.biocel.2013.05.006. Epub 2013 May 13. Int J Biochem Cell Biol. 2013. PMID: 23680674
-
P2X4 purinoceptor signaling in chronic pain.Purinergic Signal. 2012 Sep;8(3):621-8. doi: 10.1007/s11302-012-9306-7. Epub 2012 Apr 15. Purinergic Signal. 2012. PMID: 22528681 Free PMC article. Review.
-
Prostaglandin E2 as a mediator of fever: synthesis and catabolism.Front Biosci. 2004 May 1;9:1977-93. doi: 10.2741/1383. Front Biosci. 2004. PMID: 14977603 Review.
Cited by
-
Cystitis-Related Bladder Pain Involves ATP-Dependent HMGB1 Release from Macrophages and Its Downstream H2S/Cav3.2 Signaling in Mice.Cells. 2020 Jul 22;9(8):1748. doi: 10.3390/cells9081748. Cells. 2020. PMID: 32707767 Free PMC article.
-
Fluvastatin suppresses native and recombinant human P2X4 receptor function.Purinergic Signal. 2012 Jun;8(2):311-6. doi: 10.1007/s11302-011-9289-9. Epub 2012 Jan 6. Purinergic Signal. 2012. PMID: 22222818 Free PMC article.
-
Microglia P2X4R-BDNF signalling contributes to central sensitization in a recurrent nitroglycerin-induced chronic migraine model.J Headache Pain. 2020 Jan 14;21(1):4. doi: 10.1186/s10194-019-1070-4. J Headache Pain. 2020. PMID: 31937253 Free PMC article.
-
Fast Green FCF Alleviates Pain Hypersensitivity and Down-Regulates the Levels of Spinal P2X4 Expression and Pro-inflammatory Cytokines in a Rodent Inflammatory Pain Model.Front Pharmacol. 2018 May 23;9:534. doi: 10.3389/fphar.2018.00534. eCollection 2018. Front Pharmacol. 2018. PMID: 29875666 Free PMC article.
-
Inflammatory-induced spinal dorsal horn neurons hyperexcitability is mediated by P2X4 receptors.Pain Rep. 2018 May 23;3(3):e660. doi: 10.1097/PR9.0000000000000660. eCollection 2018 May. Pain Rep. 2018. PMID: 29922748 Free PMC article.
References
-
- Balboa MA, Balsinde J, Johnson CA, Dennis EA (1999) Regulation of arachidonic acid mobilization in lipopolysaccharide-activated P388D(1) macrophages by adenosine triphosphate. J Biol Chem 274: 36764–36768 - PubMed
-
- Block ML, Hong JS (2005) Microglia and inflammation-mediated neurodegeneration: multiple triggers with a common mechanism. Prog Neurobiol 76: 77–98 - PubMed
-
- Bonventre JV, Sapirstein A (2002) Group IV cytosolic phospholipase A2 (PLA2) function: insights from the knockout mouse. Adv Exp Med Biol 507: 25–31 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
