

Unique functionality of 22-nt miRNAs in triggering RDR6-dependent siRNA biogenesis from target transcripts in Arabidopsis
- PMID: 20562854
- PMCID: PMC2916640
- DOI: 10.1038/nsmb.1866
Unique functionality of 22-nt miRNAs in triggering RDR6-dependent siRNA biogenesis from target transcripts in Arabidopsis
Abstract
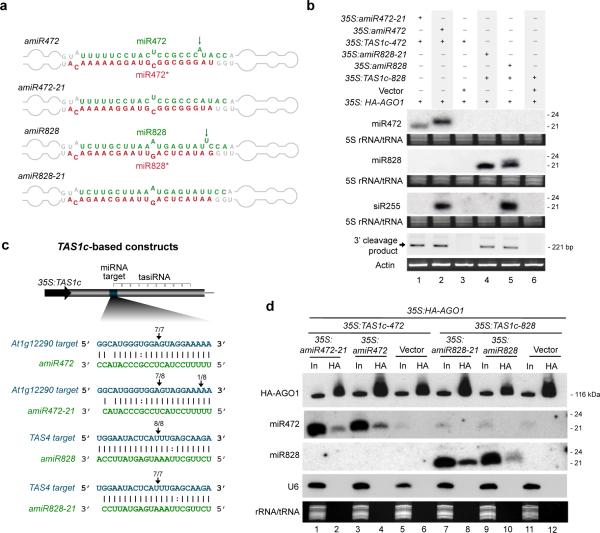
RNA interference pathways can involve amplification of secondary siRNAs by RNA-dependent RNA polymerases. In plants, RDR6-dependent secondary siRNAs arise from transcripts targeted by some microRNAs (miRNAs). Here, Arabidopsis thaliana secondary siRNAs from mRNA as well as trans-acting siRNAs are shown to be triggered through initial targeting by a 22-nucleotide (nt) miRNA that associates with AGO1. In contrast to canonical 21-nt miRNAs, 22-nt miRNAs primarily arise from foldback precursors containing asymmetric bulges. Using artificial miRNA constructs, conversion of asymmetric foldbacks to symmetric foldbacks resulted in the production of 21-nt forms of miR173, miR472 and miR828. Both 21- and 22-nt forms associated with AGO1 and guided accurate slicer activity, but only 22-nt forms were competent to trigger RDR6-dependent siRNA production from target RNA. These data suggest that AGO1 functions differentially with 21- and 22-nt miRNAs to engage the RDR6-associated amplification apparatus.
Figures



References
-
- Kim VN, Han J, Siomi MC. Biogenesis of small RNAs in animals. Nat Rev Mol Cell Biol. 2009;10:126–139. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
