

Following the intersubunit conformation of the ribosome during translation in real time
- PMID: 20562856
- PMCID: PMC4459212
- DOI: 10.1038/nsmb.1828
Following the intersubunit conformation of the ribosome during translation in real time
Abstract
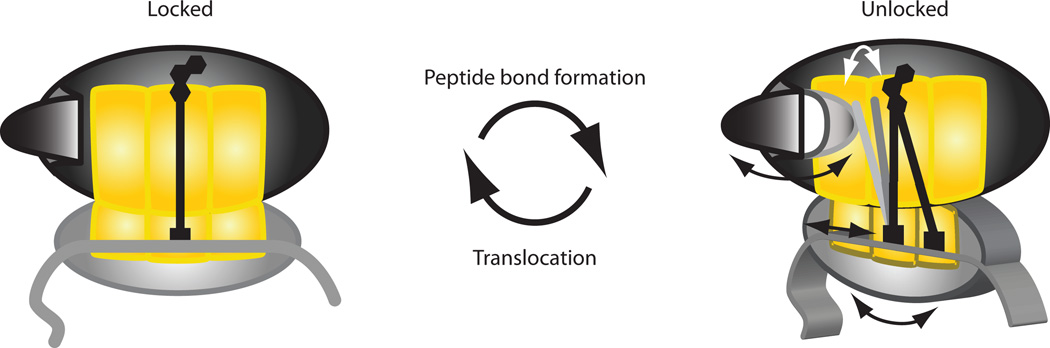
We report the direct observation of conformational rearrangements of the ribosome during multiple rounds of elongation. Using single-molecule fluorescence resonance energy transfer, we monitored the intersubunit conformation of the ribosome in real time as it proceeds from codon to codon. During each elongation cycle, the ribosome unlocks upon peptide bond formation, then reverts to the locked state upon translocation onto the next codon. Our data reveal both the specific and cumulative effects of antibiotics on individual steps of translation and uncover the processivity of the ribosome as it elongates. Our approach interrogates the precise molecular events occurring at each codon of the mRNA within the full context of ongoing translation.
Figures






References
-
- Green R, Noller HF. Ribosomes and translation. Annu. Rev. Biochem. 1997;66:679–716. - PubMed
-
- Wintermeyer W, et al. Mechanisms of elongation on the ribosome: dynamics of a macromolecular machine. Biochem Soc Trans. 2004;32:733–737. - PubMed
-
- Spirin AS. A model of the functioning ribosome: locking and unlocking of the ribosome subparticles. Cold Spring Harb Symp Quant Biol. 1969;34:197–207. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
