

Cell transformation by v-Rel reveals distinct roles of AP-1 family members in Rel/NF-kappaB oncogenesis
- PMID: 20562914
- PMCID: PMC2932816
- DOI: 10.1038/onc.2010.239
Cell transformation by v-Rel reveals distinct roles of AP-1 family members in Rel/NF-kappaB oncogenesis
Abstract
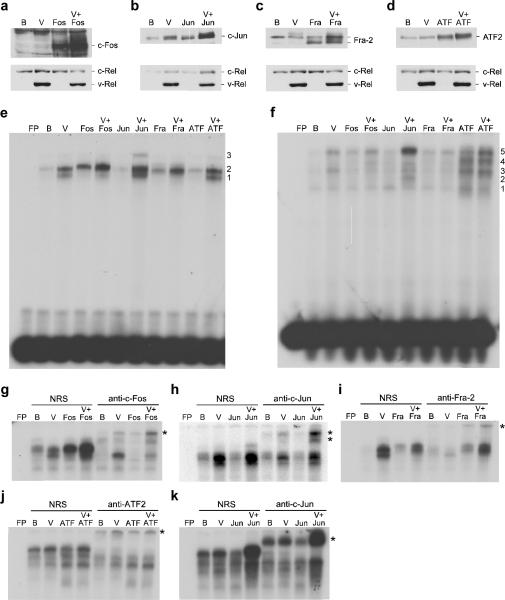
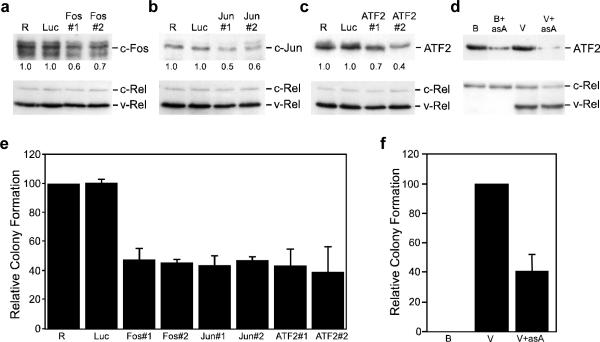
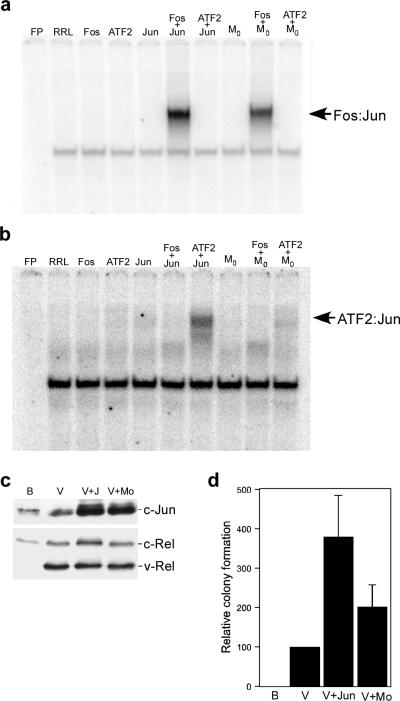
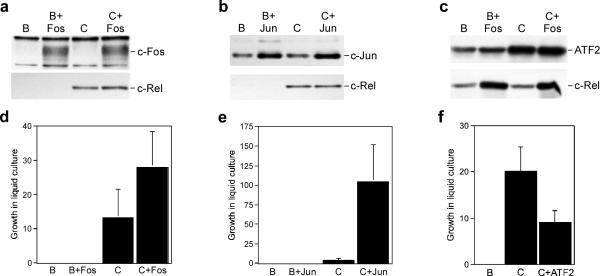
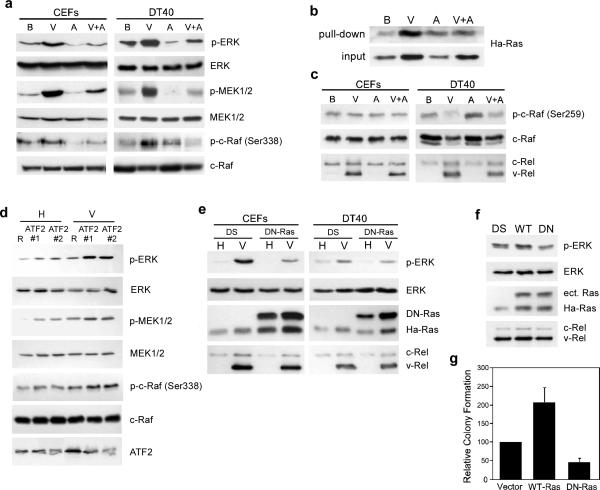
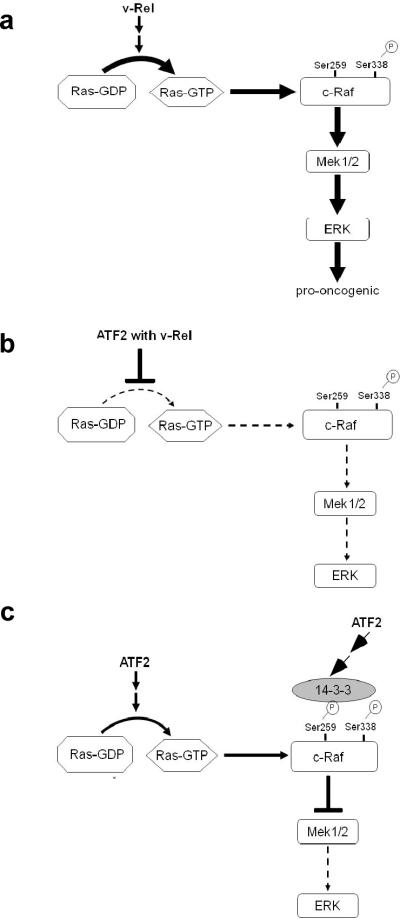
Cell transformation by the v-rel oncogene is mediated by the aberrant expression of genes that are normally tightly regulated by other Rel/NF-kappaB family members. Although a number of genes inappropriately activated or suppressed by v-Rel have been identified, their contributions to the v-Rel transformation process have been poorly characterized. Here, we examine the role of individual AP-1 proteins in v-Rel-mediated transformation. v-Rel-transformed cells exhibit elevated RNA and protein expression of c-Fos, c-Jun and ATF2 and sustained repression of Fra-2. c-Fos and c-Jun are essential in both the initiation and maintenance of v-Rel-mediated transformation, whereas Fra-2 is dispensable. By employing a c-Jun dimerization mutant, we further identified Fos/Jun heterodimers as major contributors to the v-Rel transformation process. The inability of c-Rel to induce the expression of c-Fos and c-Jun contributes to its weaker oncogenic potential relative to v-Rel. Our studies also demonstrate that v-Rel may induce AP-1 members by directly upregulating gene expression (c-fos and ATF2) and by activating pathways that stimulate AP-1 activity. Although elevated expression of ATF2 is also required for v-Rel-mediated transformation, its ectopic overexpression is inhibitory. Investigating the mode of ATF2 regulation revealed a positive feedback mechanism whereby ATF2 induces p38 MAPK phosphorylation to further induce its own activity. In addition, these studies identified Ha-Ras as an effector of v-Rel-mediated transformation and reveal a novel role for ATF2 in the inhibition of the Ras-Raf-MEK-ERK signaling pathway. Overall, these studies reveal distinct and complex roles of AP-1 proteins in Rel/NF-kappaB oncogenesis.
Figures

Similar articles
-
AP-1 factors play an important role in transformation induced by the v-rel oncogene.Mol Cell Biol. 1998 May;18(5):2997-3009. doi: 10.1128/MCB.18.5.2997. Mol Cell Biol. 1998. PMID: 9566919 Free PMC article.
-
Restriction to Fos family members of Trip6-dependent coactivation and glucocorticoid receptor-dependent trans-repression of activator protein-1.Mol Endocrinol. 2008 Aug;22(8):1767-80. doi: 10.1210/me.2007-0574. Epub 2008 Jun 5. Mol Endocrinol. 2008. PMID: 18535250 Free PMC article.
-
Expression of dominant negative Jun inhibits elevated AP-1 and NF-kappaB transactivation and suppresses anchorage independent growth of HPV immortalized human keratinocytes.Oncogene. 1998 May 28;16(21):2711-21. doi: 10.1038/sj.onc.1201798. Oncogene. 1998. PMID: 9652737
-
Distinct roles of Jun : Fos and Jun : ATF dimers in oncogenesis.Oncogene. 2001 Apr 30;20(19):2453-64. doi: 10.1038/sj.onc.1204239. Oncogene. 2001. PMID: 11402340 Review.
-
Emerging roles of ATF2 and the dynamic AP1 network in cancer.Nat Rev Cancer. 2010 Jan;10(1):65-76. doi: 10.1038/nrc2681. Nat Rev Cancer. 2010. PMID: 20029425 Free PMC article. Review.
Cited by
-
Activation of gga-miR-155 by reticuloendotheliosis virus T strain and its contribution to transformation.J Gen Virol. 2017 Apr;98(4):810-820. doi: 10.1099/jgv.0.000718. Epub 2017 Apr 28. J Gen Virol. 2017. PMID: 28113043 Free PMC article.
-
Inhibition of v-rel-Induced Oncogenesis through microRNA Targeting.Viruses. 2018 May 5;10(5):242. doi: 10.3390/v10050242. Viruses. 2018. PMID: 29734737 Free PMC article.
-
Aaptamines from the marine sponge Aaptos sp. display anticancer activities in human cancer cell lines and modulate AP-1-, NF-κB-, and p53-dependent transcriptional activity in mouse JB6 Cl41 cells.Biomed Res Int. 2014;2014:469309. doi: 10.1155/2014/469309. Epub 2014 Jul 23. Biomed Res Int. 2014. PMID: 25215281 Free PMC article.
-
ERK and JNK activation is essential for oncogenic transformation by v-Rel.Oncogene. 2010 Nov 25;29(47):6267-79. doi: 10.1038/onc.2010.359. Epub 2010 Aug 30. Oncogene. 2010. PMID: 20802521 Free PMC article.
-
Characterization of ATF2 in Rel/NFκB oncogenesis reveals its role in the regulation of Ras signaling.Small GTPases. 2011 Mar;2(2):89-94. doi: 10.4161/sgtp.2.2.15310. Small GTPases. 2011. PMID: 21776408 Free PMC article.
References
-
- Angel P, Hattori K, Smeal T, Karin M. The jun proto-oncogene is positively autoregulated by its product, Jun/AP-1. Cell. 1988;55:875–85. - PubMed
-
- Angel P, Imagawa M, Chiu R, Stein B, Imbra RJ, Rahmsdorf HJ, et al. Phorbol ester-inducible genes contain a common cis element recognized by a TPA-modulated trans-acting factor. Cell. 1987;49:729–39. - PubMed
-
- Chaudhary A, King WG, Mattaliano MD, Frost JA, Diaz B, Morrison DK, et al. Phosphatidylinositol 3-kinase regulates Raf1 through Pak phosphorylation of serine 338. Curr Biol. 2000;10:551–4. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
