

Invasive glioblastoma cells acquire stemness and increased Akt activation
- PMID: 20563248
- PMCID: PMC2887086
- DOI: 10.1593/neo.10126
Invasive glioblastoma cells acquire stemness and increased Akt activation
Abstract
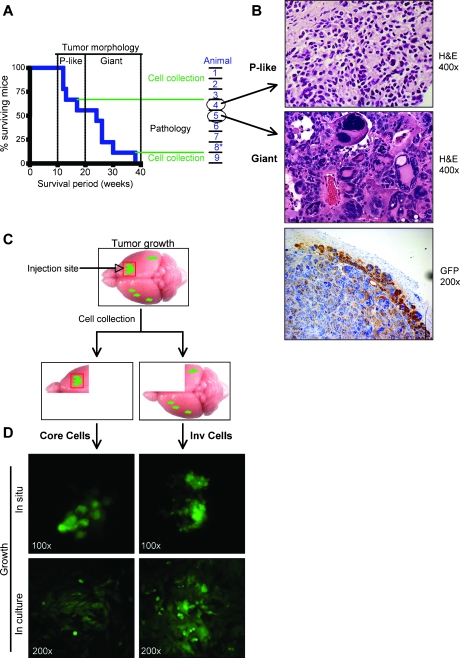
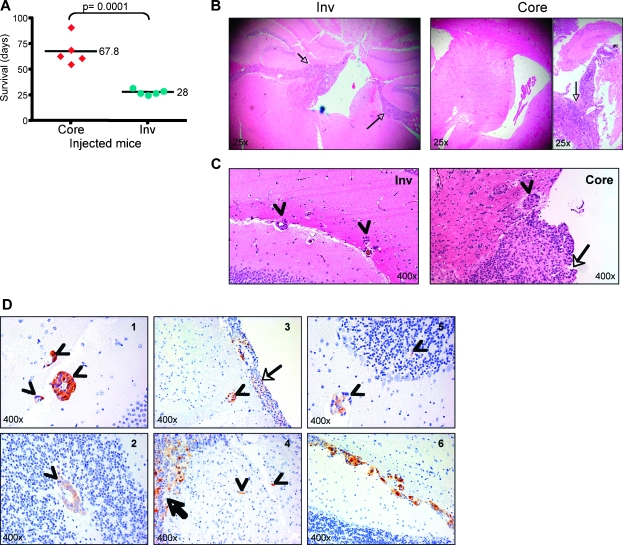
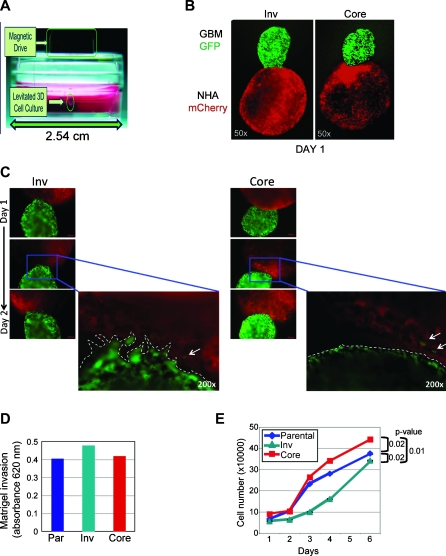
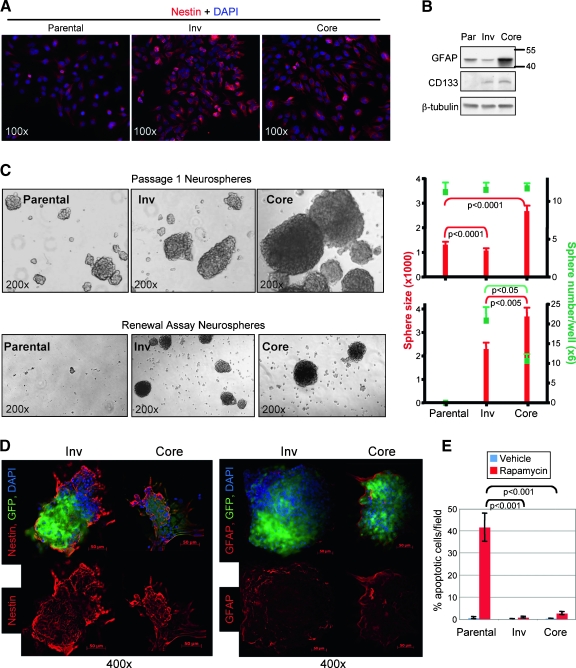
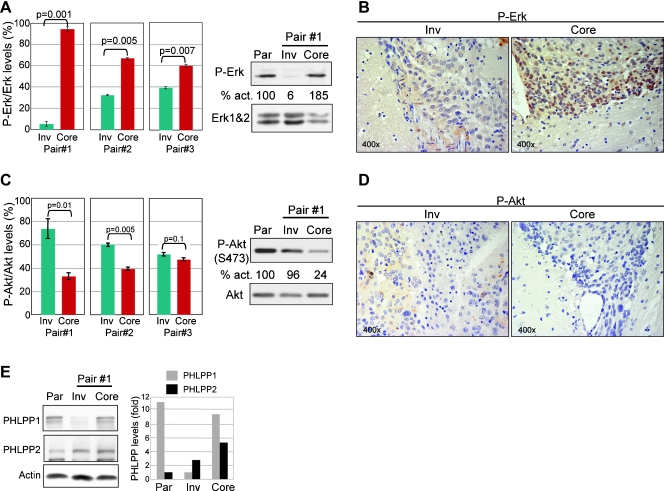
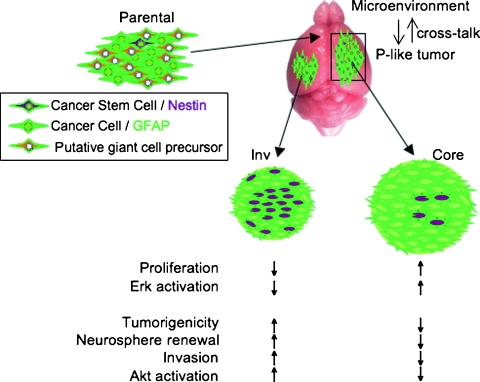
Glioblastoma multiforme (GBM) is the most frequent and most aggressive brain tumor in adults. The dismal prognosis is due to postsurgery recurrences arising from escaped invasive tumor cells. The signaling pathways activated in invasive cells are under investigation, and models are currently designed in search for therapeutic targets. We developed here an in vivo model of human invasive GBM in mouse brain from a GBM cell line with moderate tumorigenicity that allowed simultaneous primary tumor growth and dispersal of tumor cells in the brain parenchyma. This strategy allowed for the first time the isolation and characterization of matched sets of tumor mass (Core) and invasive (Inv) cells. Both cell populations, but more markedly Inv cells, acquired stem cell markers, neurosphere renewal ability, and resistance to rapamycin-induced apoptosis relative to parental cells. The comparative phenotypic analysis between Inv and Core cells showed significantly increased tumorigenicity in vivo and increased invasion with decreased proliferation in vitro for Inv cells. Examination of a large array of signaling pathways revealed extracellular signal-regulated kinase (Erk) down-modulation and Akt activation in Inv cells and an opposite profile in Core cells. Akt activation correlated with the increased tumorigenicity, stemness, and invasiveness, whereas Erk activation correlated with the proliferation of the cells. These results underscore complementary roles of the Erk and Akt pathways for GBM proliferation and dispersal and raise important implications for a concurrent inhibitory therapy.
Figures
References
-
- Furnari FB, Fenton T, Bachoo RM, Mukasa A, Stommel JM, Stegh A, Hahn WC, Ligon KL, Louis DN, Brennan C, et al. Malignant astrocytic glioma: genetics, biology, and paths to treatment. Genes Dev. 2007;21:2683–2710. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous