

Interleukin-18 as an in vivo mediator of monocyte recruitment in rodent models of rheumatoid arthritis
- PMID: 20565717
- PMCID: PMC2911912
- DOI: 10.1186/ar3055
Interleukin-18 as an in vivo mediator of monocyte recruitment in rodent models of rheumatoid arthritis
Abstract
Introduction: The function of interleukin-18 (IL-18) was investigated in pertinent animal models of rodent rheumatoid arthritis (RA) to determine its proinflammatory and monocyte recruitment properties.
Methods: We used a modified Boyden chemotaxis system to examine monocyte recruitment to recombinant human (rhu) IL-18 in vitro. Monocyte recruitment to rhuIL-18 was then tested in vivo by using an RA synovial tissue (ST) severe combined immunodeficient (SCID) mouse chimera. We defined monocyte-specific signal-transduction pathways induced by rhuIL-18 with Western blotting analysis and linked this to in vitro monocyte chemotactic activity. Finally, the ability of IL-18 to induce a cytokine cascade during acute joint inflammatory responses was examined by inducing wild-type (Wt) and IL-18 gene-knockout mice with zymosan-induced arthritis (ZIA).
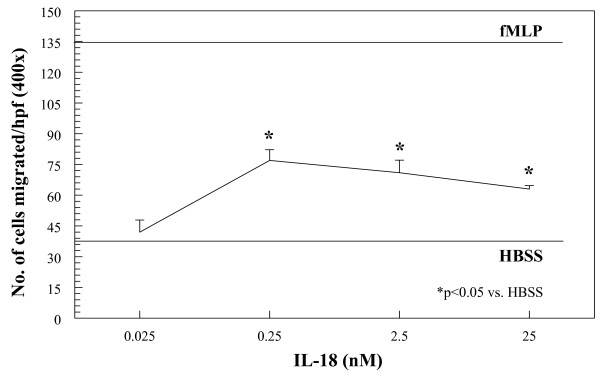
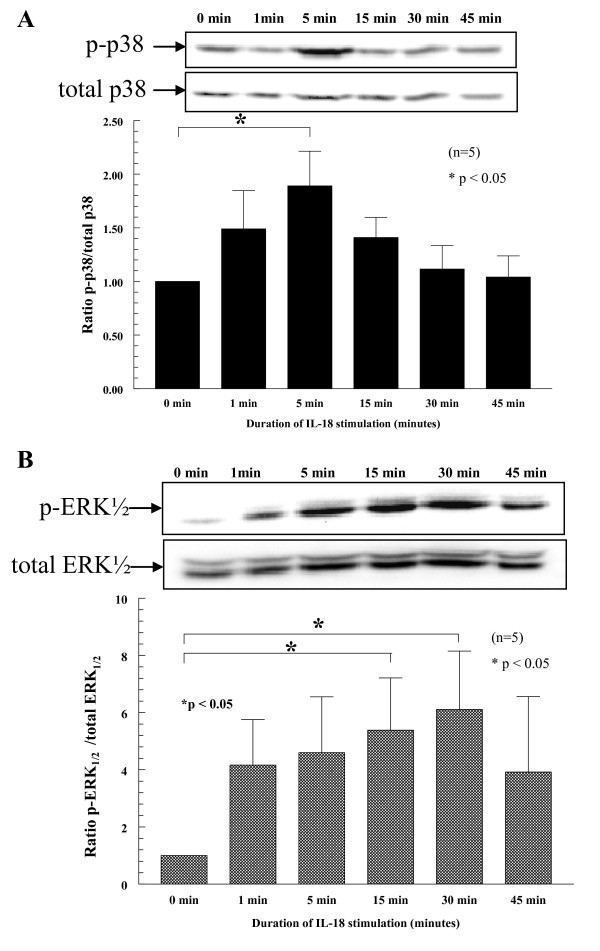
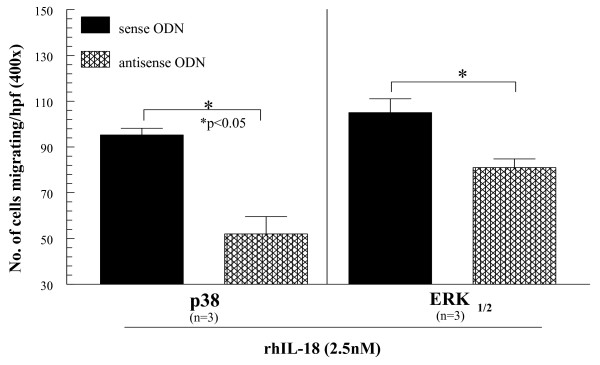
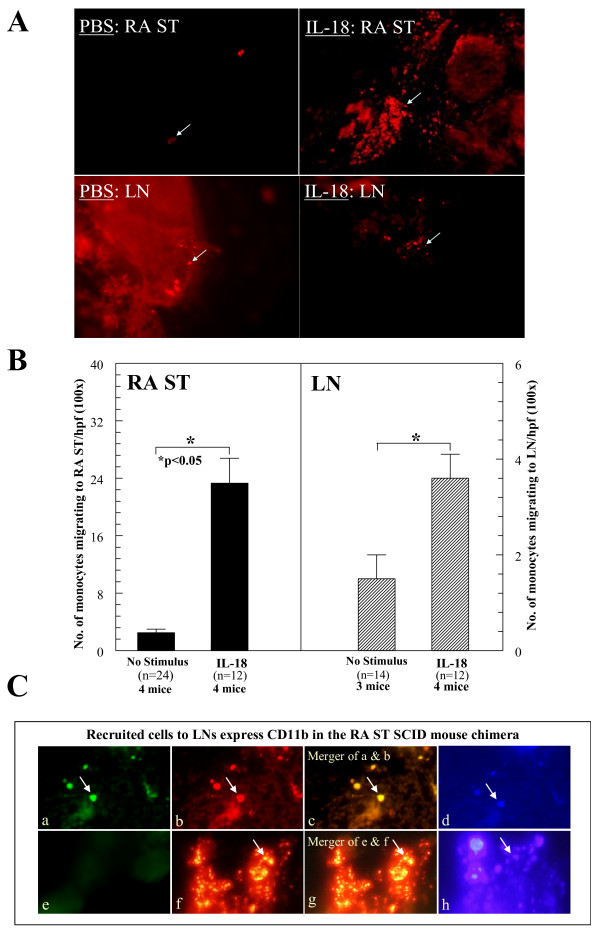
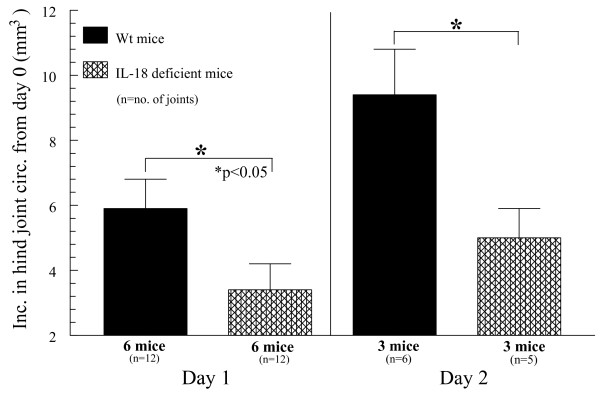
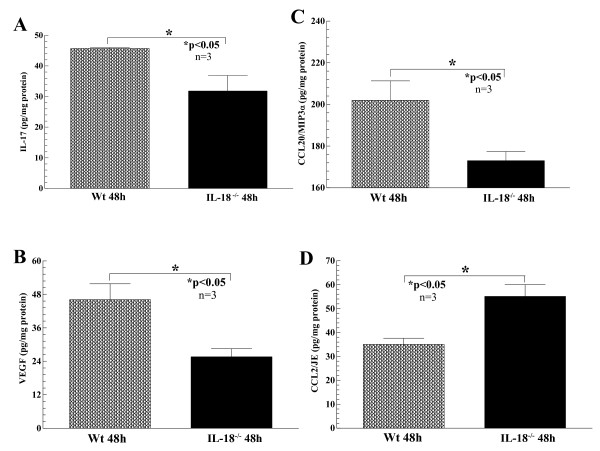
Results: We found that intragraft injected rhuIL-18 was a robust monocyte recruitment factor to both human ST and regional (inguinal) murine lymph node (LN) tissue. IL-18 gene-knockout mice also showed pronounced reductions in joint inflammation during ZIA compared with Wt mice. Many proinflammatory cytokines were reduced in IL-18 gene-knockout mouse joint homogenates during ZIA, including macrophage inflammatory protein-3alpha (MIP-3alpha/CCL20), vascular endothelial cell growth factor (VEGF), and IL-17. Signal-transduction experiments revealed that IL-18 signals through p38 and ERK1/2 in monocytes, and that IL-18-mediated in vitro monocyte chemotaxis can be significantly inhibited by disruption of this pathway.
Conclusions: Our data suggest that IL-18 may be produced in acute inflammatory responses and support the notion that IL-18 may serve a hierarchic position for initiating joint inflammatory responses.
Figures
References
-
- Boraschi D, Dinarello CA. IL-18 in autoimmunity: review. Eur Cytokine Netwk. 2006;17:224–252. - PubMed
-
- Kawashima M, Yamamura M, Taniai M, Yamauchi H, Tanimoto T, Kurimoto M, Miyawaki S, Amano T, Takeuchi T, Makino H. Levels of interleukin-18 and its binding inhibitors in the blood circulation of patients with adult-onset Still's disease. Arthritis Rheum. 2001;44:550–560. doi: 10.1002/1529-0131(200103)44:3<550::AID-ANR103>3.0.CO;2-5. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
