

doi: 10.1017/S1740925X10000098.
Epub 2010 Jun 22.
Can satellite glial cells be therapeutic targets for pain control?
Affiliations
- PMID: 20566001
- PMCID: PMC3139431
- DOI: 10.1017/S1740925X10000098
Item in Clipboard
Can satellite glial cells be therapeutic targets for pain control?
Neuron Glia Biol.
2010 Feb.
Abstract
Satellite glial cells (SGCs) undergo phenotypic changes and divide the following injury into a peripheral nerve. Nerve injury, also elicits an immune response and several antigen-presenting cells are found in close proximity to SGCs. Silencing SCG-specific molecules involved in intercellular transport (Connexin 43) or glutamate recycling (glutamine synthase) can dramatically alter nociceptive responses of normal and nerve-injured rats. Transducing SGCs with glutamic acid decarboxylase can produce analgesia in models of trigeminal pain. Taken together these data suggest that SGCs may play a role in the genesis or maintenance of pain and open a range of new possibilities for curing neuropathic pain.
Figures

Four days following CCI of the ION and 2 days after BrdU injection, BrdU immuno-labeled nuclei (red, arrows) are present in SGCs identified by the SGC-specific marker SK3 (green; Vit et al., 2006). Other BrdU positive nuclei of an unidentified population of cells are also found in close proximity to SGCs. Scale bar, A1–A3 = 50 μm; A4 = 25 μm.

Following a CCI of the ION, there is an increase in number and staining intensity of both ED2− (red cells in A1–C4) and CD45− (red cells in D1–E4) immunopositive cells. It is common to see both ED2− and CD45 immunopositive cells lying closely apposed to SGCs. In some instances the immune cells appear to lie between the neuron cell body and the SGC (B3, arrow). In some cases, SK3 positive SGCs are also immunopositive for ED2 (C1–C4, arrow) or CD45 (E1–E4, arrow). In other cases, cells that appear to be SGCs based on morphological and location criteria (F1–G2), exhibit both ED2 and CD45 immunoreactivity. Letter a and b indicate the same element in F1–F3. N = nucleus. Scale bars, A1–A2 = 120 μm; B1–B4 = 10 μm; D1–D2 = 60 μm; E1–E4 = 30 μm; F1–F3 = 50 μm; G1–G2 = 20 μm.

(A) Cx43 immunostaining (green) is present throughout the trigeminal ganglia as a diffuse cytoplasmic staining together with many small brightly fluorescent puncta. (B) Following a CCI of the ION, there is a marked increase in both cytoplasmic staining and number of immunofluorescent puncta. The increase was confirmed by Western blot quantification (Cx43 expression density; contralateral side 7.3 ± 3.33; ipsilateral side 35.33 ± 2.66; P < 0.001. n = 8 per group). (C) Two days after injection of Cx43 dsRNA into the trigeminal ganglia on the side of the CCI, there is a marked reduction in Cx43 immunostaining to levels less than in uninjured animals. DiI (red) injected with the Cx43 dsRNA to confirm the injection site is indicated by the arrow. (D) When tested in the operant training apparatus there is a reduction in drinking following CCI of the ION (interrupted line). Between 1 and 3 days following Cx43 dsRNA injection the number of licks returns to baseline levels indicating analgesia on the operated side. * < 0.05, ** < 0.001 compared to baseline. # < 0.05, ## < 0.001 compared to Globin dsRNA control. n = 6. Scale bar, 30 μm.
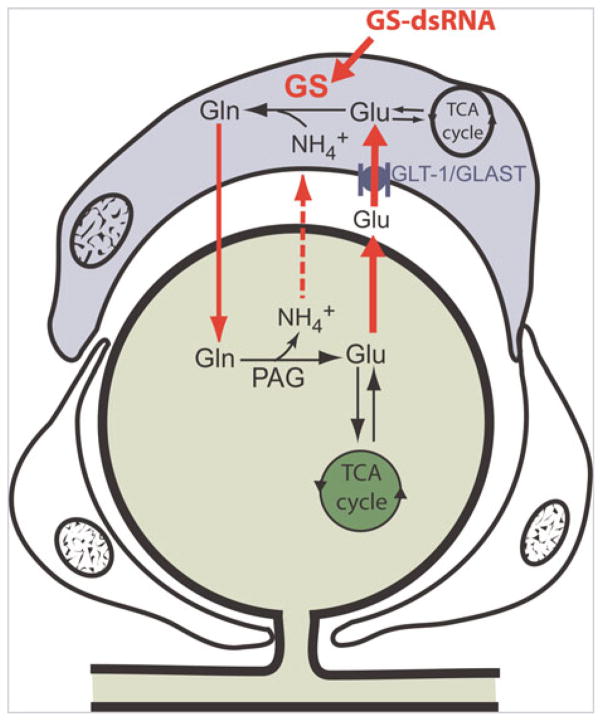
The glial glutamate transporter (GLAST) transports extracellular glutamate (Glu) into the SGC where glutamine synthetase (GS) converts the glutamate to glutamine (Gln) which is eventually recycled to the neuron for conversion into glutamate. Silencing GS using dsRNA-mediated RNAi will reduce the amount of glutamine available to neurons for conversion to glutamate.

(A) GS immunoreactivity (green) is present in SGCs of the trigeminal ganglion. (B) Three days following injection of GS dsRNA into the trigeminal ganglion, GS immunoreactivity is greatly reduced. The arrow indicates DiI injected with the dsRNA. (C) Orofacial formalin tests were performed four days after the injection of GS dsRNA or control dsRNA. Compared to naive and control dsRNA rats, GS dsRNA resulted in significant decrease in pain behavior that occurred in the second phase of the formalin test. *P < 0.05, **P < 0.01, ***P < 0.001. n = 8. Scale bar, 30 μm.
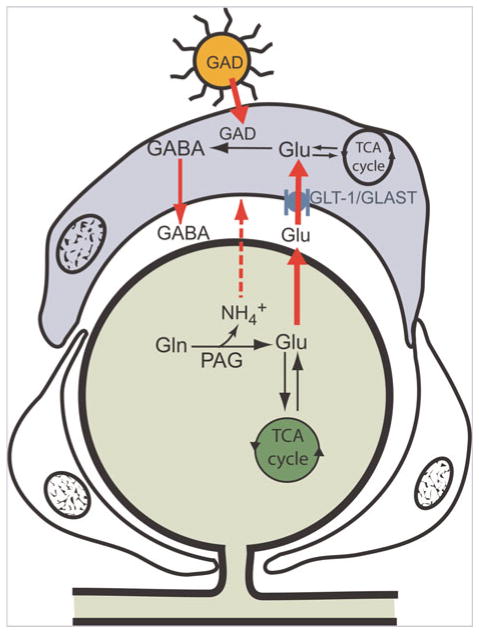
Glutamate taken up by SGCs through the GLAST transporter is normally converted to glutamine via GS (see Fig. 4). Introducing the GAD65 transgene into the SGC via an adenovirus (AdGAD65) provides the pathway for the glutamate to be converted to GABA, which is then released by bulk transport to inhibit nearby primary sensory neurons by acting on GABAA receptors.

(A) After AdGAD65 introduction into the trigeminal ganglion, GAD65 immunostaining (green) is found in SGCs (e.g. arrow) identified by GLAST immunostaining (red). (B) Six days after injection of AdGAD65, there is a significant reduction of the nociceptive response in second phase of the orofacial formalin compared to rats injected with a control virus (AdGFP) or naïve rats. AdGAD65 versus naïve: **P < 0.01, ***P < 0.001; AdGAD65 versus AdGFP: *P < 0.05, **P < 0.01, ***P < 0.001. n = 8. Bar = 30 μm.
References
-
- Adler JE, Nico L, VandeVord P, Skoff AM. Modulation of neuropathic pain by a glial-derived factor. Pain Medicine. 2009;10:1229–1236. - PubMed
-
- Bhargava A, Dallman MF, Pearce D, Choi S. Long double-stranded RNA-mediated RNA interference as a tool to achieve site-specific silencing of hypothalamic neuropeptides. Brain Research and Brain Research Protocol. 2004;13:115–125. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
