

The ribonucleotide reductase inhibitor, Sml1, is sequentially phosphorylated, ubiquitylated and degraded in response to DNA damage
- PMID: 20566477
- PMCID: PMC2965251
- DOI: 10.1093/nar/gkq552
The ribonucleotide reductase inhibitor, Sml1, is sequentially phosphorylated, ubiquitylated and degraded in response to DNA damage
Abstract
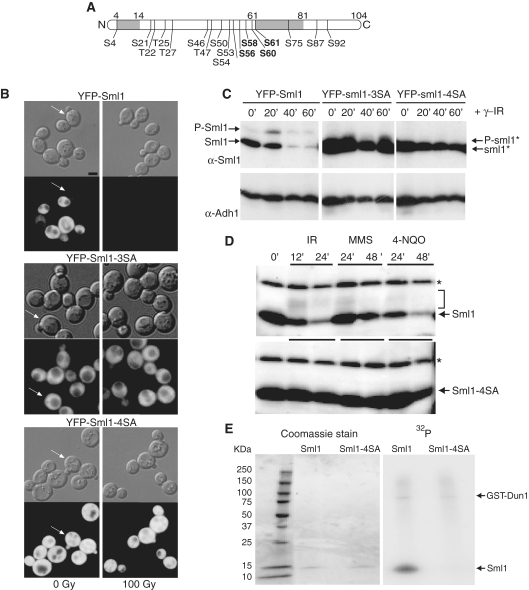
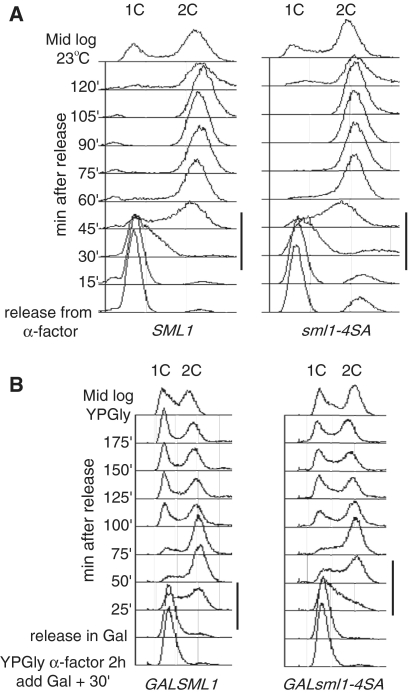
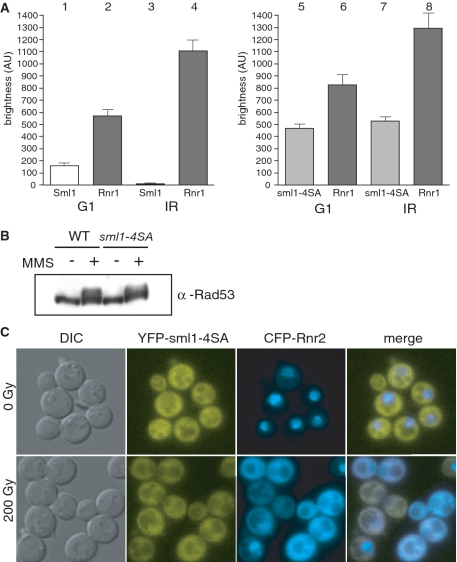
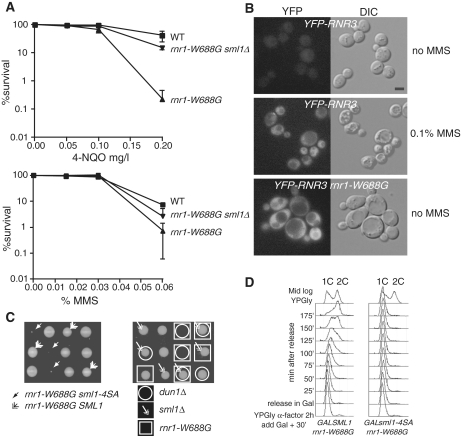
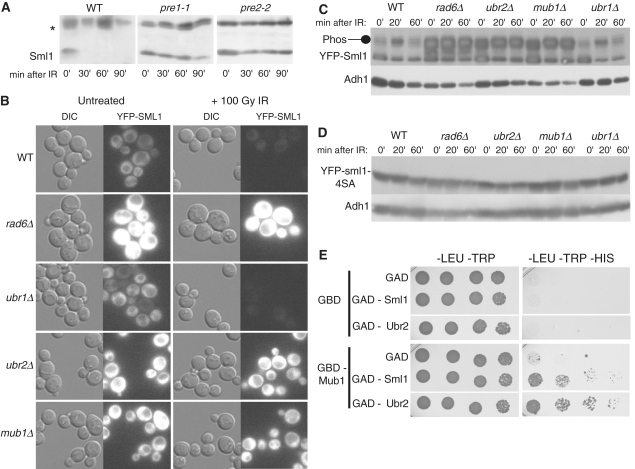
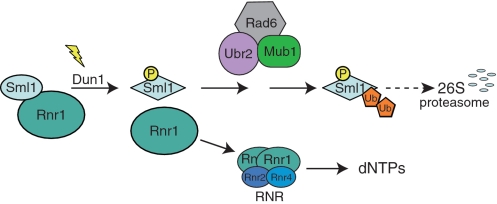
Regulation of ribonucleotide reductase (RNR) is important for cell survival and genome integrity in the face of genotoxic stress. The Mec1/Rad53/Dun1 DNA damage response kinase cascade exhibits multifaceted controls over RNR activity including the regulation of the RNR inhibitor, Sml1. After DNA damage, Sml1 is degraded leading to the up-regulation of dNTP pools by RNR. Here, we probe the requirements for Sml1 degradation and identify several sites required for in vivo phosphorylation and degradation of Sml1 in response to DNA damage. Further, in a strain containing a mutation in Rnr1, rnr1-W688G, mutation of these sites in Sml1 causes lethality. Degradation of Sml1 is dependent on the 26S proteasome. We also show that degradation of phosphorylated Sml1 is dependent on the E2 ubiquitin-conjugating enzyme, Rad6, the E3 ubiquitin ligase, Ubr2, and the E2/E3-interacting protein, Mub1, which form a complex previously only implicated in the ubiquitylation of Rpn4.
Figures
References
-
- Chabes A, Georgieva B, Domkin V, Zhao X, Rothstein R, Thelander L. Survival of DNA damage in yeast directly depends on increased dNTP levels allowed by relaxed feedback inhibition of ribonucleotide reductase. Cell. 2003;112:391–401. - PubMed
-
- Reichard P. Interactions between deoxyribonucleotide and DNA synthesis. Annu. Rev. Biochem. 1988;57:349–374. - PubMed
-
- Elledge SJ, Davis RW. Two genes differentially regulated in the cell cycle and by DNA-damaging agents encode alternative regulatory subunits of ribonucleotide reductase. Genes Dev. 1990;4:740–751. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
