

Dissecting the involvement of tropomyosin-related kinase A and p75 neurotrophin receptor signaling in NGF deficit-induced neurodegeneration
- PMID: 20566851
- PMCID: PMC2901482
- DOI: 10.1073/pnas.1007181107
Dissecting the involvement of tropomyosin-related kinase A and p75 neurotrophin receptor signaling in NGF deficit-induced neurodegeneration
Abstract
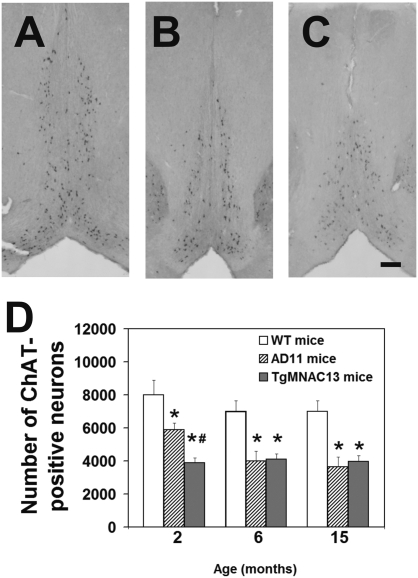
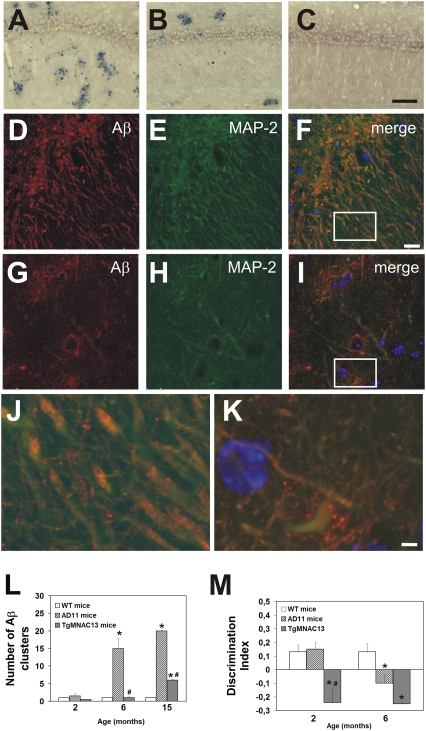
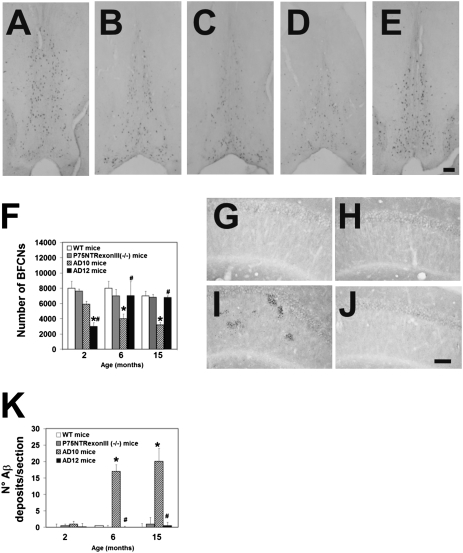
NGF, the principal neurotrophic factor for basal forebrain cholinergic neurons (BFCNs), has been correlated to Alzheimer's disease (AD) because of the selective vulnerability of BFCNs in AD. These correlative links do not substantiate a comprehensive cause-effect mechanism connecting NGF deficit to overall AD neurodegeneration. A demonstration that neutralizing NGF activity could have consequences beyond a direct interference with the cholinergic system came from studies in the AD11 mouse model, in which the expression of a highly specific anti-NGF antibody determines a neurodegeneration that encompasses several features of human AD. Because the transgenic antibody binds to mature NGF much more strongly than to proNGF and prevents binding of mature NGF to the tropomyosin-related kinase A (TrkA) receptor and to p75 neurotrophin receptor (p75NTR), we postulated that neurodegeneration in AD11 mice is provoked by an imbalance of proNGF/NGF signaling and, consequently, of TrkA/p75NTR signaling. To test this hypothesis, in this study we characterize the phenotype of two lines of transgenic mice, one in which TrkA signaling is inhibited by neutralizing anti-TrkA antibodies and a second one in which anti-NGF mice were crossed to p75NTR(exonIII(-/-)) mice to abrogate p75NTR signaling. TrkA neutralization determines a strong cholinergic deficit and the appearance of beta-amyloid peptide (Abeta) but no tau-related pathology. In contrast, abrogating p75NTR signaling determines a full rescue of the cholinergic and Abeta phenotype of anti-NGF mice, but tau hyperphosphorylation is exacerbated. Thus, we demonstrate that inhibiting TrkA signaling activates Abeta accumulation and that different streams of AD neurodegeneration are related in complex ways to TrkA versus p75NTR signaling.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Nerve growth factor survival signaling in cultured hippocampal neurons is mediated through TrkA and requires the common neurotrophin receptor P75.Neuroscience. 2002;115(4):1089-108. doi: 10.1016/s0306-4522(02)00539-0. Neuroscience. 2002. PMID: 12453482
-
The Intersection of NGF/TrkA Signaling and Amyloid Precursor Protein Processing in Alzheimer's Disease Neuropathology.Int J Mol Sci. 2017 Jun 20;18(6):1319. doi: 10.3390/ijms18061319. Int J Mol Sci. 2017. PMID: 28632177 Free PMC article. Review.
-
Type I interferons up-regulate the expression and signalling of p75 NTR/TrkA receptor complex in differentiated human SH-SY5Y neuroblastoma cells.Neuropharmacology. 2014 Apr;79:321-34. doi: 10.1016/j.neuropharm.2013.12.002. Epub 2013 Dec 11. Neuropharmacology. 2014. PMID: 24333329
-
A TrkA-to-p75NTR molecular switch activates amyloid beta-peptide generation during aging.Biochem J. 2005 Oct 1;391(Pt 1):59-67. doi: 10.1042/BJ20050700. Biochem J. 2005. PMID: 15966860 Free PMC article.
-
ProNGF and Neurodegeneration in Alzheimer's Disease.Front Neurosci. 2019 Feb 22;13:129. doi: 10.3389/fnins.2019.00129. eCollection 2019. Front Neurosci. 2019. PMID: 30853882 Free PMC article. Review.
Cited by
-
Immunosympathectomy as the first phenotypic knockout with antibodies.Proc Natl Acad Sci U S A. 2013 Mar 26;110(13):4877-85. doi: 10.1073/pnas.1217586110. Epub 2013 Mar 20. Proc Natl Acad Sci U S A. 2013. PMID: 23515328 Free PMC article.
-
p75NTR ectodomain is a physiological neuroprotective molecule against amyloid-beta toxicity in the brain of Alzheimer's disease.Mol Psychiatry. 2015 Nov;20(11):1301-10. doi: 10.1038/mp.2015.49. Epub 2015 Apr 28. Mol Psychiatry. 2015. PMID: 25917367 Free PMC article.
-
proNGF Measurement in Cerebrospinal Fluid Samples of a Large Cohort of Living Patients With Alzheimer's Disease by a New Automated Immunoassay.Front Aging Neurosci. 2021 Oct 27;13:741414. doi: 10.3389/fnagi.2021.741414. eCollection 2021. Front Aging Neurosci. 2021. PMID: 34776928 Free PMC article.
-
Single cycle structure-based humanization of an anti-nerve growth factor therapeutic antibody.PLoS One. 2012;7(3):e32212. doi: 10.1371/journal.pone.0032212. Epub 2012 Mar 5. PLoS One. 2012. PMID: 22403636 Free PMC article.
-
Nerve growth factor regulates axial rotation during early stages of chick embryo development.Proc Natl Acad Sci U S A. 2012 Feb 7;109(6):2009-14. doi: 10.1073/pnas.1121138109. Epub 2012 Jan 25. Proc Natl Acad Sci U S A. 2012. PMID: 22308471 Free PMC article.
References
-
- Levi-Montalcini R. Effects of mouse tumor transplantation on the nervous system. Ann N Y Acad Sci. 1952;55:330–344. - PubMed
-
- Salehi A, Delcroix JD, Mobley WC. Traffic at the intersection of neurotrophic factor signaling and neurodegeneration. Trends Neurosci. 2003;26:73–80. - PubMed
-
- Cattaneo A, Capsoni S, Paoletti F. Towards noninvasive Nerve Growth Factor therapies for Alzheimer's disease. J Alzheimers Dis. 2008;15:255–283. - PubMed
-
- Bruno MA, et al. Amyloid beta-induced nerve growth factor dysmetabolism in Alzheimer disease. J Neuropathol Exp Neurol. 2009;68:857–869. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
