

Adenosine-to-inosine RNA editing shapes transcriptome diversity in primates
- PMID: 20566853
- PMCID: PMC2901480
- DOI: 10.1073/pnas.1006183107
Adenosine-to-inosine RNA editing shapes transcriptome diversity in primates
Abstract
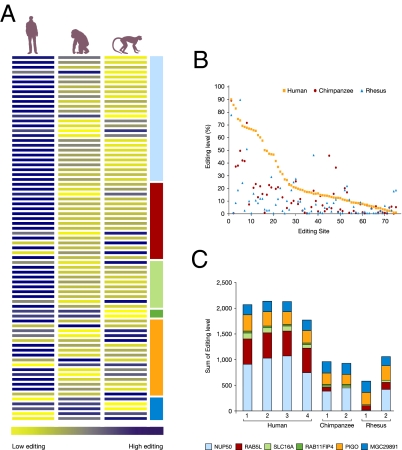
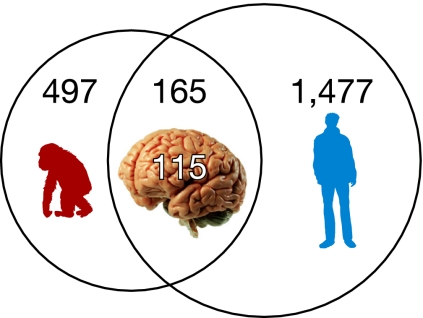
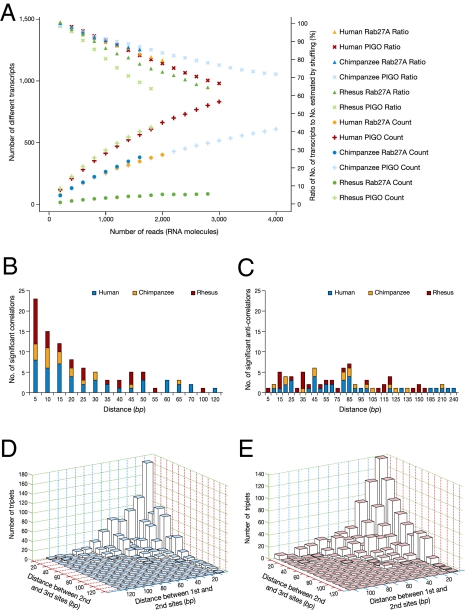
Human and chimpanzee genomes are almost identical, yet humans express higher brain capabilities. Deciphering the basis for this superiority is a long sought-after challenge. Adenosine-to-inosine (A-to-I) RNA editing is a widespread modification of the transcriptome. The editing level in humans is significantly higher compared with nonprimates, due to exceptional editing within the primate-specific Alu sequences, but the global editing level of nonhuman primates has not been studied so far. Here we report the sequencing of transcribed Alu sequences in humans, chimpanzees, and rhesus monkeys. We found that, on average, the editing level in the transcripts analyzed is higher in human brain compared with nonhuman primates, even where the genomic Alu structure is unmodified. Correlated editing is observed for pairs and triplets of specific adenosines along the Alu sequences. Moreover, new editable species-specific Alu insertions, subsequent to the human-chimpanzee split, are significantly enriched in genes related to neuronal functions and neurological diseases. The enhanced editing level in the human brain and the association with neuronal functions both hint at the possible contribution of A-to-I editing to the development of higher brain function. We show here that combinatorial editing is the most significant contributor to the transcriptome repertoire and suggest that Alu editing adapted by natural selection may therefore serve as an alternate information mechanism based on the binary A/I code.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
