

Disruption of the olivo-cerebellar circuit by Purkinje neuron-specific ablation of BK channels
- PMID: 20566869
- PMCID: PMC2901450
- DOI: 10.1073/pnas.1001745107
Disruption of the olivo-cerebellar circuit by Purkinje neuron-specific ablation of BK channels
Abstract
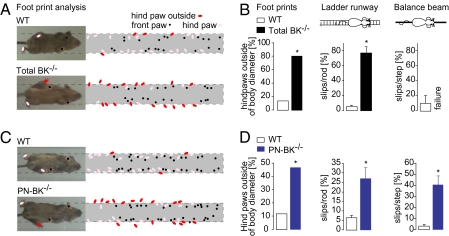
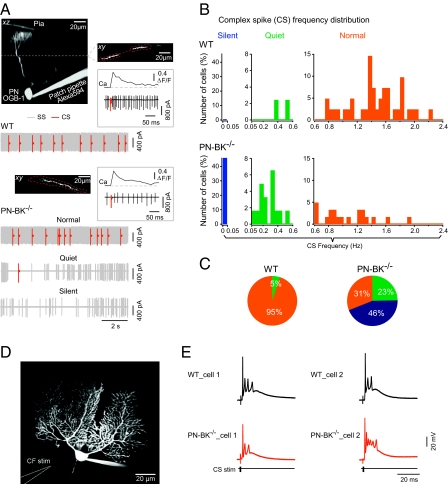
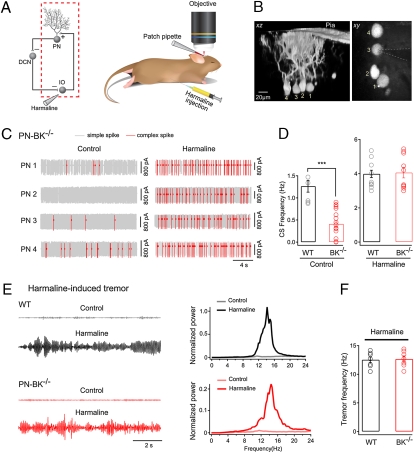
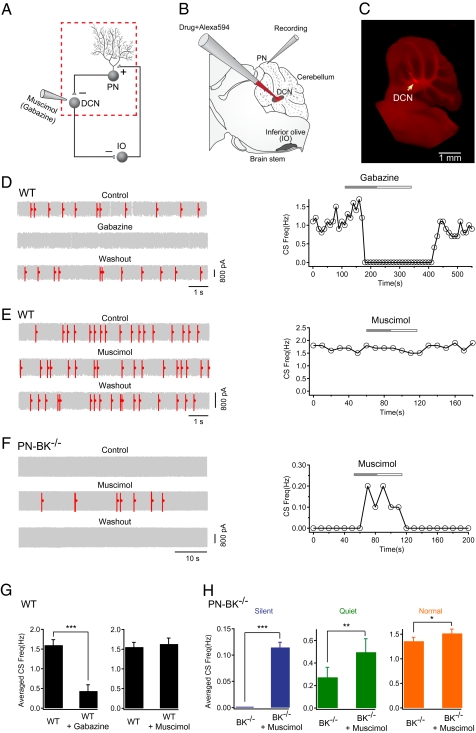
The large-conductance voltage- and calcium-activated potassium (BK) channels are ubiquitously expressed in the brain and play an important role in the regulation of neuronal excitation. Previous work has shown that the total deletion of these channels causes an impaired motor behavior, consistent with a cerebellar dysfunction. Cellular analyses showed that a decrease in spike firing rate occurred in at least two types of cerebellar neurons, namely in Purkinje neurons (PNs) and in Golgi cells. To determine the relative role of PNs, we developed a cell-selective mouse mutant, which lacked functional BK channels exclusively in PNs. The behavioral analysis of these mice revealed clear symptoms of ataxia, indicating that the BK channels of PNs are of major importance for normal motor coordination. By using combined two-photon imaging and patch-clamp recordings in these mutant mice, we observed a unique type of synaptic dysfunction in vivo, namely a severe silencing of the climbing fiber-evoked complex spike activity. By performing targeted pharmacological manipulations combined with simultaneous patch-clamp recordings in PNs, we obtained direct evidence that this silencing of climbing fiber activity is due to a malfunction of the tripartite olivo-cerebellar feedback loop, consisting of the inhibitory synaptic connection of PNs to the deep cerebellar nuclei (DCN), followed by a projection of inhibitory DCN afferents to the inferior olive, the origin of climbing fibers. Taken together, our results establish an essential role of BK channels of PNs for both cerebellar motor coordination and feedback regulation in the olivo-cerebellar loop.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
