

Plasticity of postsynaptic, but not presynaptic, GABAB receptors in SSADH deficient mice
- PMID: 20570675
- PMCID: PMC3654531
- DOI: 10.1016/j.expneurol.2010.05.022
Plasticity of postsynaptic, but not presynaptic, GABAB receptors in SSADH deficient mice
Abstract
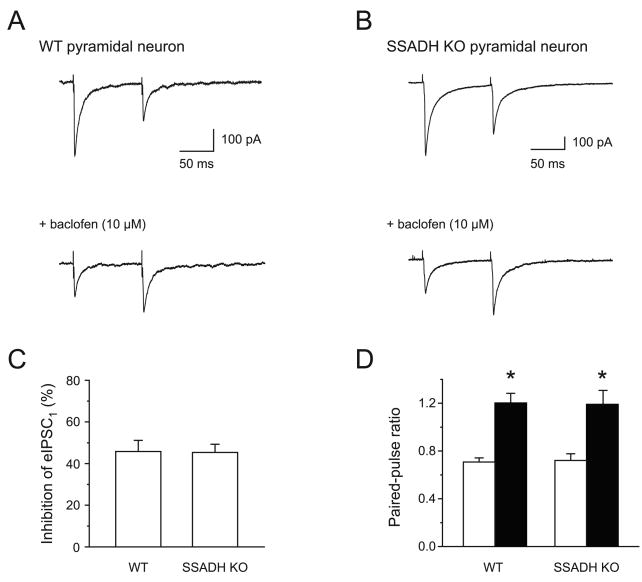
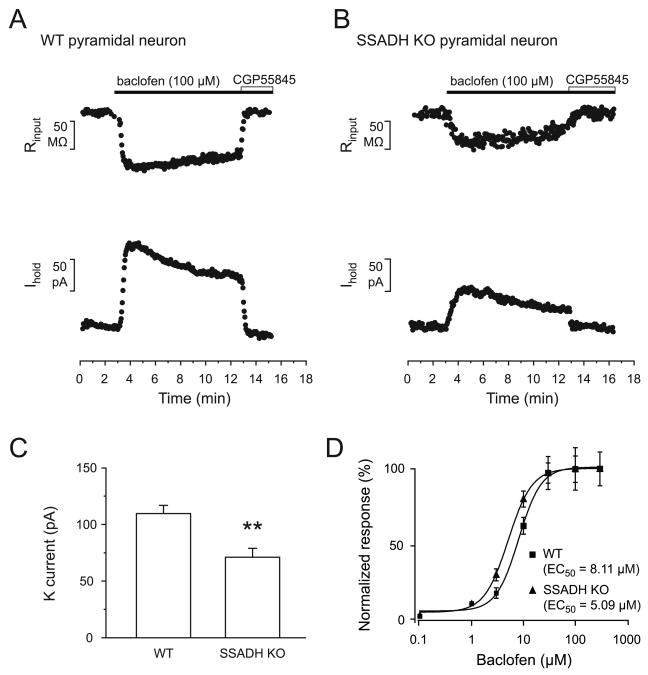
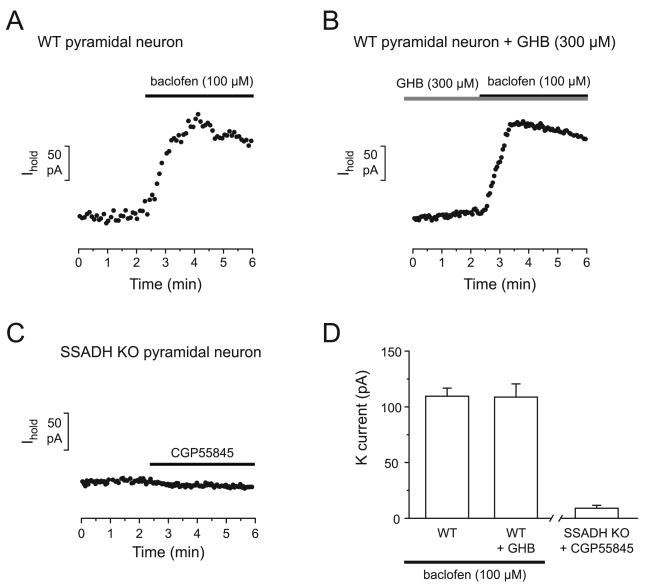
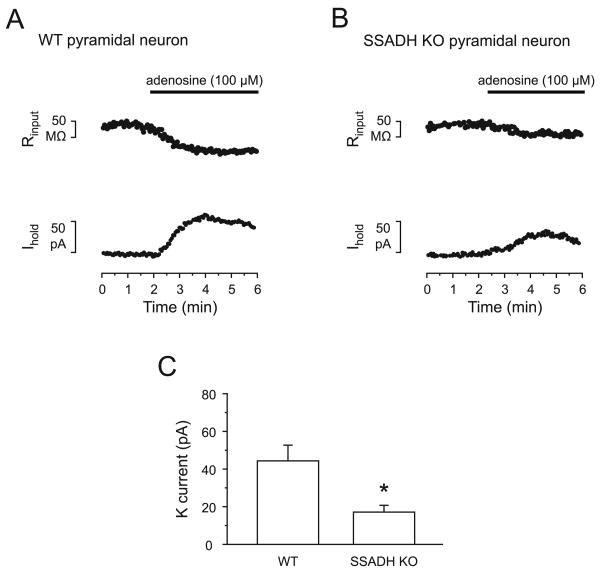
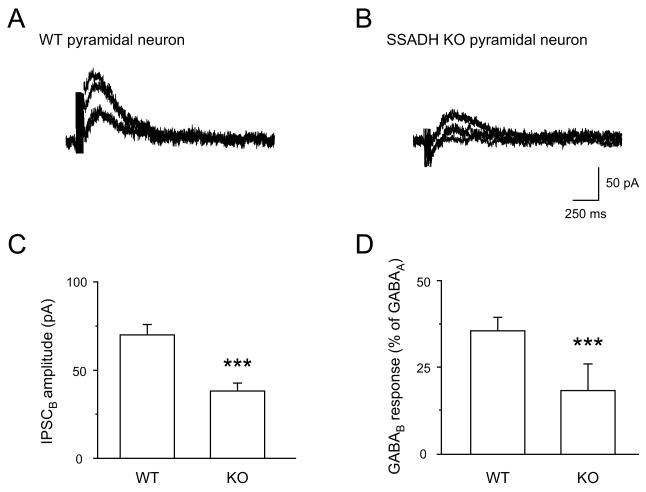
Succinic semialdehyde dehydrogenase (SSADH) deficiency is an autosomal-recessively inherited disorder of gamma-aminobutyrate (GABA) catabolism characterized by ataxia and epilepsy. Since SSADH is responsible for GABA break-down downstream of GABA transaminase, patients manifest high extracellular levels of GABA, as well as the GABA(B) receptor (GABA(B)R) agonist gamma-hydroxybutyrate (GHB). SSADH knockout (KO) mice display absence seizures, which progress into lethal tonic-clonic seizures at around 3weeks of age. It is hypothesized that desensitization of GABA(B)Rs plays an important role in the disease, although detailed studies of pre- and postsynaptic GABA(B)Rs are not available. We performed patch-clamp recordings from layer 2/3 pyramidal neurons in neocortical brain slices of wild-type (WT) and SSADH KO mice. Electrical stimulation of GABAergic fibers during wash in of the GABA(B)R agonist baclofen revealed no difference in presynaptic GABA(B)R mediated inhibition of GABA release between WT and SSADH KO mice. In contrast, a significant decrease in postsynaptic baclofen-induced potassium currents was seen in SSADH KO mice. This reduction was unlikely to be caused by accumulation of potassium, GABA or GHB in the brain slices, or an altered expression of regulators of G-protein signaling (RGS) proteins. Finally, adenosine-induced potassium currents were also reduced in SSADH KO mice, which could suggest heterologous desensitization of the G-protein dependent effectors, leading to a reduction in G-protein coupled inwardly rectifying potassium (GIRK) channel responses. Our findings indicate that high GABA and GHB levels desensitize postsynaptic, but not certain presynaptic, GABA(B)Rs, promoting a decrease in GIRK channel function. These changes could contribute to the development of seizures in SSADH KO mice and potentially also in affected patients.
Copyright 2010 Elsevier Inc. All rights reserved.
Figures
Similar articles
-
Succinic semialdehyde dehydrogenase deficiency: GABAB receptor-mediated function.Brain Res. 2006 May 23;1090(1):15-22. doi: 10.1016/j.brainres.2006.02.131. Epub 2006 May 2. Brain Res. 2006. PMID: 16647690
-
Plasticity of pre- and postsynaptic GABAB receptor function in the paraventricular nucleus in spontaneously hypertensive rats.Am J Physiol Heart Circ Physiol. 2008 Aug;295(2):H807-15. doi: 10.1152/ajpheart.00259.2008. Epub 2008 Jun 20. Am J Physiol Heart Circ Physiol. 2008. PMID: 18567709 Free PMC article.
-
Effect of gene dosage on single-cell hippocampal electrophysiology in a murine model of SSADH deficiency (gamma-hydroxybutyric aciduria).Epilepsy Res. 2010 Jun;90(1-2):39-46. doi: 10.1016/j.eplepsyres.2010.03.005. Epub 2010 Apr 3. Epilepsy Res. 2010. PMID: 20363598 Free PMC article.
-
Comparative genomics of aldehyde dehydrogenase 5a1 (succinate semialdehyde dehydrogenase) and accumulation of gamma-hydroxybutyrate associated with its deficiency.Hum Genomics. 2009 Jan;3(2):106-20. doi: 10.1186/1479-7364-3-2-106. Hum Genomics. 2009. PMID: 19164088 Free PMC article. Review.
-
Murine succinate semialdehyde dehydrogenase deficiency.Ann Neurol. 2003;54 Suppl 6:S81-90. doi: 10.1002/ana.10625. Ann Neurol. 2003. PMID: 12891658 Review.
Cited by
-
mTOR inhibitors rescue premature lethality and attenuate dysregulation of GABAergic/glutamatergic transcription in murine succinate semialdehyde dehydrogenase deficiency (SSADHD), a disorder of GABA metabolism.J Inherit Metab Dis. 2016 Nov;39(6):877-886. doi: 10.1007/s10545-016-9959-4. Epub 2016 Aug 12. J Inherit Metab Dis. 2016. PMID: 27518770 Free PMC article.
-
Effectiveness of nicotinic agonists as desensitizers at presynaptic α4β2- and α4α5β2-nicotinic acetylcholine receptors.Nicotine Tob Res. 2014 Mar;16(3):297-305. doi: 10.1093/ntr/ntt146. Epub 2013 Sep 19. Nicotine Tob Res. 2014. PMID: 24052501 Free PMC article.
-
Succinic semialdehyde dehydrogenase deficiency (SSADHD): Pathophysiological complexity and multifactorial trait associations in a rare monogenic disorder of GABA metabolism.Neurochem Int. 2016 Oct;99:72-84. doi: 10.1016/j.neuint.2016.06.009. Epub 2016 Jun 14. Neurochem Int. 2016. PMID: 27311541 Free PMC article. Review.
-
GABAB-ergic motor cortex dysfunction in SSADH deficiency.Neurology. 2012 Jul 3;79(1):47-54. doi: 10.1212/WNL.0b013e31825dcf71. Epub 2012 Jun 20. Neurology. 2012. PMID: 22722631 Free PMC article.
-
Multiple inhibitory G-protein-coupled receptors resist acute desensitization in the presynaptic but not postsynaptic compartments of neurons.J Neurosci. 2012 Jul 25;32(30):10192-200. doi: 10.1523/JNEUROSCI.1227-12.2012. J Neurosci. 2012. PMID: 22836254 Free PMC article.
References
-
- Bettler B, Kaupmann K, Mosbacher J, Gassmann M. Molecular structure and physiological functions of GABA(B) receptors. Physiol Rev. 2004;84:835–867. - PubMed
-
- Bischoff S, Leonhard S, Reymann N, Schuler V, Shigemoto R, Kaupmann K, Bettler B. Spatial distribution of GABA(B)R1 receptor mRNA and binding sites in the rat brain. J Comp Neurol. 1999;412:1–16. - PubMed
-
- Bowery NG. GABA(B) receptor: a site of therapeutic benefit. Curr Opin Pharmacol. 2006;6:37–43. - PubMed
-
- Brown JT, Gill CH, Farmer CE, Lanneau C, Randall AD, Pangalos MN, Collingridge GL, Davies CH. Mechanisms contributing to the exacerbated epileptiform activity in hippocampal slices of GABAB1 receptor subunit knockout mice. Epilepsy Res. 2003;57:121–136. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
