

Review
doi: 10.1016/j.ymeth.2010.06.003.
Epub 2010 Jun 4.
What lessons can be learned from studying the folding of homologous proteins?
Affiliations
- PMID: 20570731
- PMCID: PMC2965948
- DOI: 10.1016/j.ymeth.2010.06.003
Item in Clipboard
Review
What lessons can be learned from studying the folding of homologous proteins?
Methods.
2010 Sep.
Abstract
The studies of the folding of structurally related proteins have proved to be a very important tool for investigating protein folding. Here we review some of the insights that have been gained from such studies. Our highlighted studies show just how such an investigation should be designed and emphasise the importance of the synergy between experiment and theory. We also stress the importance of choosing the right system carefully, exploiting the excellent structural and sequence databases at our disposal.
Copyright (c) 2010 Elsevier Inc. All rights reserved.
Figures
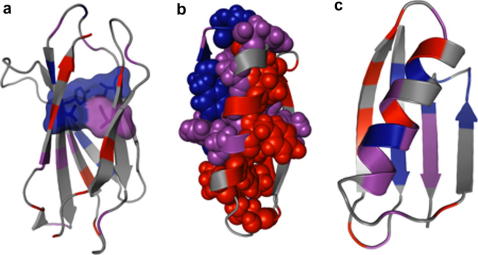
Example Φ-value patterns and how they illustrate aspects of the folding pathway. In each case the Φ-values are mapped onto a cartoon representation of the native state, with high values coloured blue, moderate values in magenta and low values in red. (a) TNfn3 : the highest Φ-values are predominantly found in the central β-strands, and decrease away from a common-core ring of interactions, known as the ‘key-residues’, which set up the complex Greek-key topology of the native state. (b) Protein L : the symmetry of the native state is completely broken during folding, with a high degree of structure in the C-terminal β-hairpin but virtually no structure formation in the N-terminal hairpin. (c) Protein G : in contrast to Protein L, this transition state exhibits significant structure in the first β-hairpin, and little structure formation in the C-terminal hairpin.
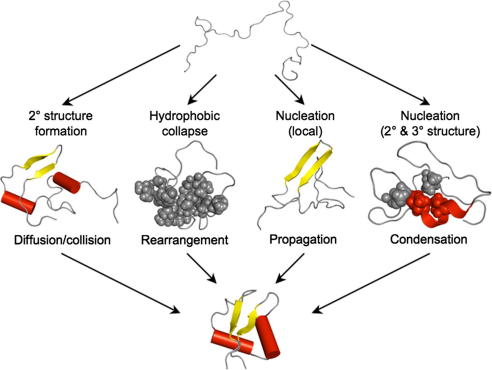
A description of the four ‘classical’ folding mechanisms. (1) The framework model suggests that local elements of secondary structure form first. These then diffuse together, collide and adhere to produce the correct tertiary structure in the rate determining step. (2) The hydrophobic collapse model implies that a protein collapses rapidly around its hydrophobic side-chains, and then rearranges from the restricted conformation of this ‘molten-globule’ intermediate. (3) The nucleation propagation model states that local interactions form a small amount of native secondary structure, which acts as a nucleus for the outward propagation of further native structure. (4) The nucleation condensation model suggests the presence of a metastable nucleus that is unable to trigger folding until a sufficient number of stabilising long-range interactions have built up. Once this occurs, the native structure condenses so rapidly that the nucleus is not yet fully formed in the transition state.
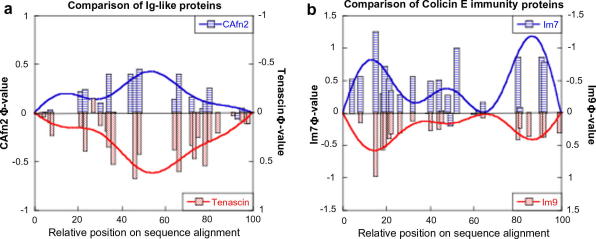
Comparison of homologous proteins. (a) Folding of two Ig-like domains, CAfn2 and TNfn3, both members of the fnIII superfamily. The pattern of Φ-values is extremely well conserved between the two domains implying equivalent folding pathways . (b) Comparison of the folding of two colicin E immunity proteins. The pattern of Φ-values is extremely well conserved between the two proteins, despite the fact that Im9 folds with two-state kinetics whereas Im7 populates an on-pathway folding intermediate .
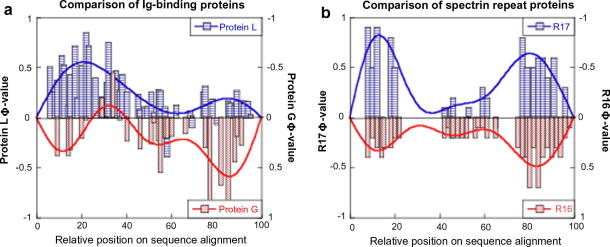
Comparison of the folding of homologous proteins with topologically symmetric native states. (a) Proteins L and G are both Ig-binding domains and exhibit a ‘breakdown of symmetry’ in their transition state ensembles. The pattern of Φ-values for Protein L is essentially a mirror-image of that found for Protein G . (b) R16 and R17 are both spectrin repeat domains and fold through a transition state that is almost symmetrical. The pattern of Φ-values is well conserved between the two domains .
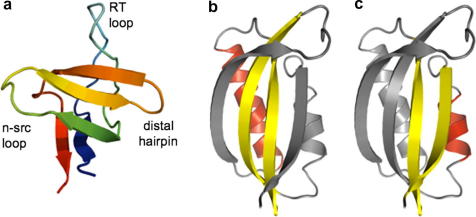
Examples of proteins studied by circular permutation. (a) α-Spectrin SH3-domain: cutting the RT loop does not result in a change in transition state structure (centred around the 3-stranded central sheet and the distal hairpin and n-src loop). Cleaving the distal hairpin, however, results in a different, more diffuse nucleus . (b and c) Two different nucleation motifs within the ferredoxin-like fold. (b) Wild-type S6T and U1A predominantly fold using a nucleus comprising the two central β-strands and the longer α1 helix . (c) S6A, ADA2h and circular permutants of S6T fold using an alternative two-strand-helix nucleation motif .
Similar articles
-
Crystal structure of the MYB domain of the RAD transcription factor from Antirrhinum majus.Proteins. 2006 Dec 1;65(4):1041-5. doi: 10.1002/prot.21136. Proteins. 2006. PMID: 17044043 No abstract available.
-
Unifying features in protein-folding mechanisms.Proc Natl Acad Sci U S A. 2003 Nov 11;100(23):13286-91. doi: 10.1073/pnas.1835776100. Epub 2003 Oct 31. Proc Natl Acad Sci U S A. 2003. PMID: 14595026 Free PMC article.
-
Insights into the folding pathway of the Engrailed Homeodomain protein using replica exchange molecular dynamics simulations.J Mol Graph Model. 2010 Nov;29(3):481-91. doi: 10.1016/j.jmgm.2010.09.007. Epub 2010 Oct 8. J Mol Graph Model. 2010. PMID: 21035367
-
Ultrafast and downhill protein folding.Curr Opin Struct Biol. 2007 Feb;17(1):38-47. doi: 10.1016/j.sbi.2007.01.001. Epub 2007 Jan 12. Curr Opin Struct Biol. 2007. PMID: 17223539 Review.
-
Protein folding and unfolding at atomic resolution.Cell. 2002 Feb 22;108(4):573-82. doi: 10.1016/s0092-8674(02)00620-7. Cell. 2002. PMID: 11909527 Review.
Cited by
-
Protein unfolding rates correlate as strongly as folding rates with native structure.Protein Sci. 2015 Apr;24(4):580-7. doi: 10.1002/pro.2606. Epub 2014 Dec 26. Protein Sci. 2015. PMID: 25422093 Free PMC article.
-
Folding and binding pathways of BH3-only proteins are encoded within their intrinsically disordered sequence, not templated by partner proteins.J Biol Chem. 2018 Jun 22;293(25):9718-9723. doi: 10.1074/jbc.RA118.002791. Epub 2018 May 1. J Biol Chem. 2018. PMID: 29716994 Free PMC article.
-
Mechanism of assembly of the non-covalent spectrin tetramerization domain from intrinsically disordered partners.J Mol Biol. 2014 Jan 9;426(1):21-35. doi: 10.1016/j.jmb.2013.08.027. Epub 2013 Sep 17. J Mol Biol. 2014. PMID: 24055379 Free PMC article.
-
Rationalizing the Role of Monosodium Glutamate in the Protein Aggregation Through Biophysical Approaches: Potential Impact on Neurodegeneration.Front Neurosci. 2021 Mar 4;15:636454. doi: 10.3389/fnins.2021.636454. eCollection 2021. Front Neurosci. 2021. PMID: 33746704 Free PMC article. Review.
-
Energetic frustrations in protein folding at residue resolution: a homologous simulation study of Im9 proteins.PLoS One. 2014 Jan 31;9(1):e87719. doi: 10.1371/journal.pone.0087719. eCollection 2014. PLoS One. 2014. PMID: 24498176 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
