

Deficiency of chemokine receptor CCR1 causes osteopenia due to impaired functions of osteoclasts and osteoblasts
- PMID: 20571024
- PMCID: PMC2937910
- DOI: 10.1074/jbc.M109.099424
Deficiency of chemokine receptor CCR1 causes osteopenia due to impaired functions of osteoclasts and osteoblasts
Abstract
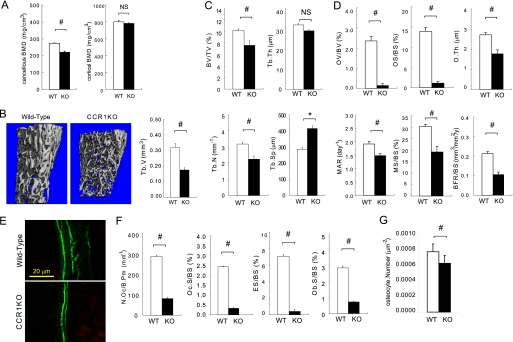
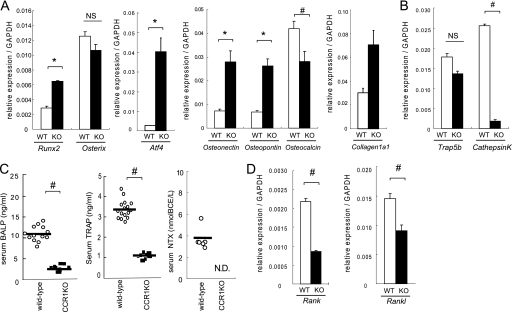
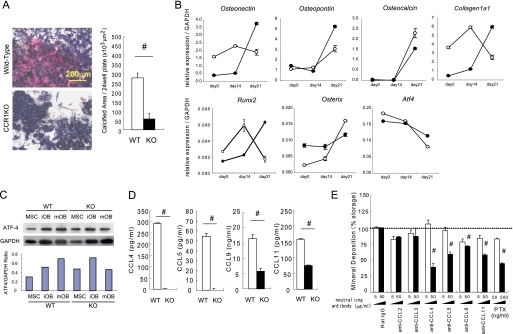
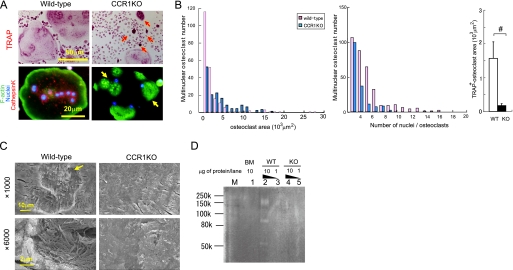
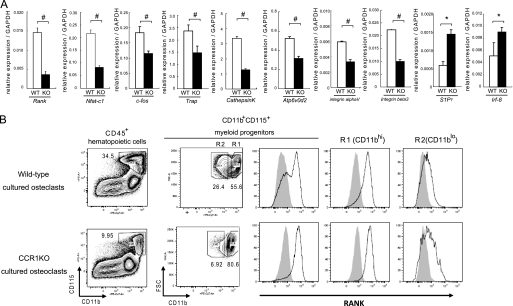
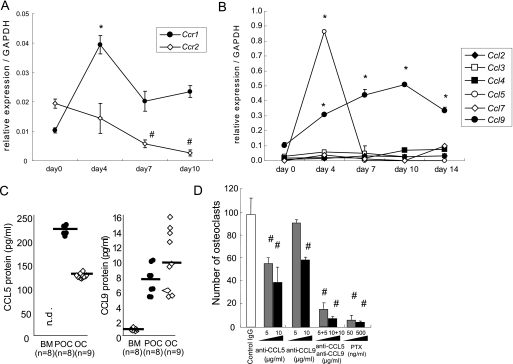
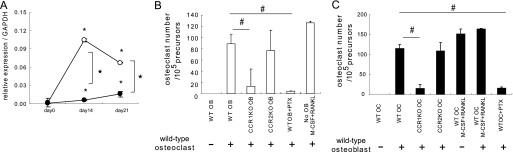
Chemokines are characterized by the homing activity of leukocytes to targeted inflammation sites. Recent research indicates that chemokines play more divergent roles in various phases of pathogenesis as well as immune reactions. The chemokine receptor, CCR1, and its ligands are thought to be involved in inflammatory bone destruction, but their physiological roles in the bone metabolism in vivo have not yet been elucidated. In the present study, we investigated the roles of CCR1 in bone metabolism using CCR1-deficient mice. Ccr1(-/-) mice have fewer and thinner trabecular bones and low mineral bone density in cancellous bones. The lack of CCR1 affects the differentiation and function of osteoblasts. Runx2, Atf4, Osteopontin, and Osteonectin were significantly up-regulated in Ccr1(-/-) mice despite sustained expression of Osterix and reduced expression of Osteocalcin, suggesting a lower potential for differentiation into mature osteoblasts. In addition, mineralized nodule formation was markedly disrupted in cultured osteoblastic cells isolated from Ccr1(-/-) mice. Osteoclastogenesis induced from cultured Ccr1(-/-) bone marrow cells yielded fewer and smaller osteoclasts due to the abrogated cell-fusion. Ccr1(-/-) osteoclasts exerted no osteolytic activity concomitant with reduced expressions of Rank and its downstream targets, implying that the defective osteoclastogenesis is involved in the bone phenotype in Ccr1(-/-) mice. The co-culture of wild-type osteoclast precursors with Ccr1(-/-) osteoblasts failed to facilitate osteoclastogenesis. This finding is most likely due to a reduction in Rankl expression. These observations suggest that the axis of CCR1 and its ligands are likely to be involved in cross-talk between osteoclasts and osteoblasts by modulating the RANK-RANKL-mediated interaction.
Figures
References
-
- Charo I. F., Ransohoff R. M. (2006) N. Engl. J. Med. 354, 610–621 - PubMed
-
- Oba Y., Lee J. W., Ehrlich L. A., Chung H. Y., Jelinek D. F., Callander N. S., Horuk R., Choi S. J., Roodman G. D. (2005) Exp. Hematol. 33, 272–278 - PubMed
-
- Kim M. S., Magno C. L., Day C. J., Morrison N. A. (2006) J. Cell Biochem. 97, 512–518 - PubMed
-
- Menu E., De Leenheer E., De Raeve H., Coulton L., Imanishi T., Miyashita K., Van Valckenborgh E., Van Riet I., Van Camp B., Horuk R., Croucher P., Vanderkerken K. (2006) Clin. Exp. Metastasis 23, 291–300 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
