

Regulation of freezing tolerance and flowering in temperate cereals: the VRN-1 connection
- PMID: 20571115
- PMCID: PMC2923912
- DOI: 10.1104/pp.110.159079
Regulation of freezing tolerance and flowering in temperate cereals: the VRN-1 connection
Abstract
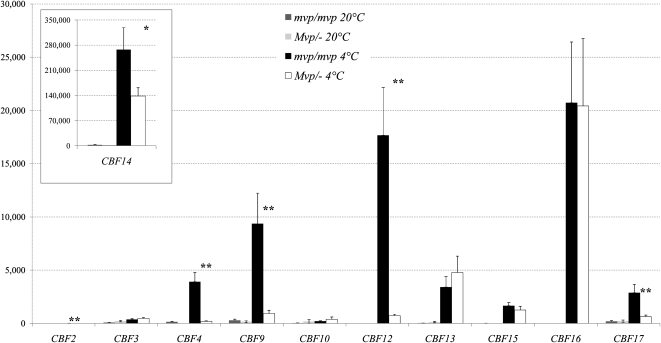
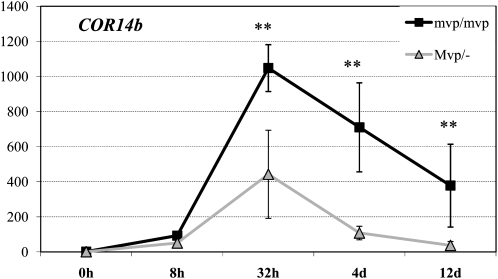
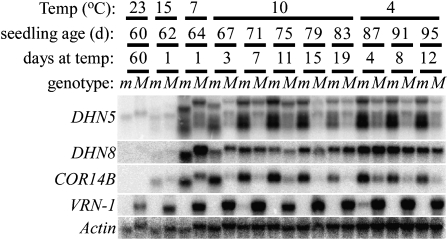
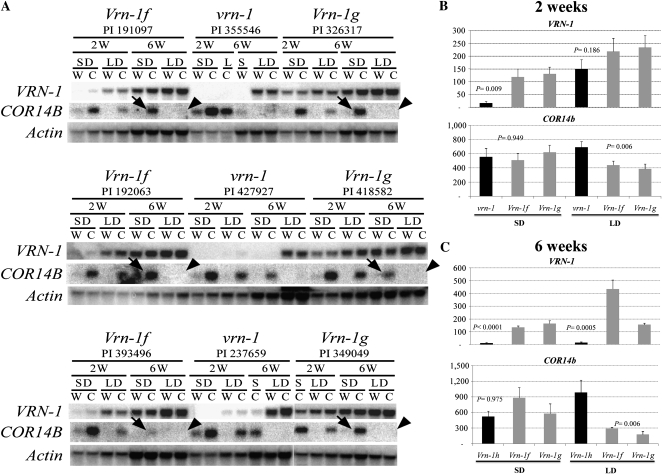
In winter wheat (Triticum spp.) and barley (Hordeum vulgare) varieties, long exposures to nonfreezing cold temperatures accelerate flowering time (vernalization) and improve freezing tolerance (cold acclimation). However, when plants initiate their reproductive development, freezing tolerance decreases, suggesting a connection between the two processes. To better understand this connection, we used two diploid wheat (Triticum monococcum) mutants, maintained vegetative phase (mvp), that carry deletions encompassing VRN-1, the major vernalization gene in temperate cereals. Homozygous mvp/mvp plants never flower, whereas plants carrying at least one functional VRN-1 copy (Mvp/-) exhibit normal flowering and high transcript levels of VRN-1 under long days. The Mvp/- plants showed reduced freezing tolerance and reduced transcript levels of several cold-induced C-REPEAT BINDING FACTOR transcription factors and COLD REGULATED genes (COR) relative to the mvp/mvp plants. Diploid wheat accessions with mutations in the VRN-1 promoter, resulting in high transcript levels under both long and short days, showed a significant down-regulation of COR14b under long days but not under short days. Taken together, these studies suggest that VRN-1 is required for the initiation of the regulatory cascade that down-regulates the cold acclimation pathway but that additional genes regulated by long days are required for the down-regulation of the COR genes. In addition, our results show that allelic variation in VRN-1 is sufficient to determine differences in freezing tolerance, suggesting that quantitative trait loci for freezing tolerance previously mapped on this chromosome region are likely a pleiotropic effect of VRN-1 rather than the effect of a separate closely linked locus (FROST RESISTANCE-1), as proposed in early freezing tolerance studies.
Figures
Similar articles
-
Regulation of VRN-1 vernalization genes in normal and transgenic polyploid wheat.Plant Physiol. 2005 Aug;138(4):2364-73. doi: 10.1104/pp.105.064287. Epub 2005 Jul 29. Plant Physiol. 2005. PMID: 16055679 Free PMC article.
-
Copy number and haplotype variation at the VRN-A1 and central FR-A2 loci are associated with frost tolerance in hexaploid wheat.Theor Appl Genet. 2014 May;127(5):1183-97. doi: 10.1007/s00122-014-2290-2. Epub 2014 Mar 14. Theor Appl Genet. 2014. PMID: 24626953 Free PMC article.
-
The effect of day-neutral mutations in barley and wheat on the interaction between photoperiod and vernalization.Theor Appl Genet. 2013 Sep;126(9):2267-77. doi: 10.1007/s00122-013-2133-6. Epub 2013 Jun 5. Theor Appl Genet. 2013. PMID: 23737074 Free PMC article.
-
Allelic Variations in Vernalization (Vrn) Genes in Triticum spp.Genes (Basel). 2024 Feb 17;15(2):251. doi: 10.3390/genes15020251. Genes (Basel). 2024. PMID: 38397240 Free PMC article. Review.
-
Expression of dehydrins in wheat and barley under different temperatures.Plant Sci. 2011 Jan;180(1):46-52. doi: 10.1016/j.plantsci.2010.07.003. Epub 2010 Jul 14. Plant Sci. 2011. PMID: 21421346 Review.
Cited by
-
Application of image-based phenotyping tools to identify QTL for in-field winter survival of winter wheat (Triticum aestivum L.).Theor Appl Genet. 2019 Sep;132(9):2591-2604. doi: 10.1007/s00122-019-03373-6. Epub 2019 Jun 8. Theor Appl Genet. 2019. PMID: 31177292
-
QTL analyses and comparative genetic mapping of frost tolerance, winter survival and drought tolerance in meadow fescue (Festuca pratensis Huds.).Theor Appl Genet. 2011 Aug;123(3):369-82. doi: 10.1007/s00122-011-1590-z. Epub 2011 Apr 20. Theor Appl Genet. 2011. PMID: 21505831
-
Comparative analysis of the cold acclimation and freezing tolerance capacities of seven diploid Brachypodium distachyon accessions.Ann Bot. 2014 Mar;113(4):681-93. doi: 10.1093/aob/mct283. Epub 2013 Dec 8. Ann Bot. 2014. PMID: 24323247 Free PMC article.
-
A Multi-Environment Trial Analysis of Frost Susceptibility in Wheat and Barley Under Australian Frost-Prone Field Conditions.Front Plant Sci. 2021 Aug 19;12:722637. doi: 10.3389/fpls.2021.722637. eCollection 2021. Front Plant Sci. 2021. PMID: 34490019 Free PMC article.
-
Relationship Between Dehydrin Accumulation and Winter Survival in Winter Wheat and Barley Grown in the Field.Front Plant Sci. 2019 Jan 29;10:7. doi: 10.3389/fpls.2019.00007. eCollection 2019. Front Plant Sci. 2019. PMID: 30761163 Free PMC article.
References
-
- Abe M, Kobayashi Y, Yamamoto S, Daimon Y, Yamaguchi A, Ikeda Y, Ichinoki H, Notaguchi M, Goto K, Araki T. (2005) FD, a bZIP protein mediating signals from the floral pathway integrator FT at the shoot apex. Science 309: 1052–1056 - PubMed
-
- Båga M, Chodaparambil SV, Limin AE, Pecar M, Fowler DB, Chibbar RN. (2007) Identification of quantitative trait loci and associated candidate genes for low-temperature tolerance in cold-hardy winter wheat. Funct Integr Genomics 7: 53–68 - PubMed
-
- Brule-Babel AL, Fowler DB. (1988) Genetic control of cold hardiness and vernalization requirement in winter wheat. Crop Sci 28: 879–884
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
