

Review
doi: 10.3109/10409238.2010.496772.
Ribonucleoprotein multimers and their functions
Affiliations
- PMID: 20572804
- PMCID: PMC2939948
- DOI: 10.3109/10409238.2010.496772
Item in Clipboard
Review
Ribonucleoprotein multimers and their functions
Crit Rev Biochem Mol Biol.
2010 Oct.
Abstract
Ribonucleoproteins (RNPs) play key roles in many cellular processes and often function as RNP enzymes. Similar to proteins, some of these RNPs exist and function as multimers, either homomeric or heteromeric. While in some cases the mechanistic function of multimerization is well understood, the functional consequences of multimerization of other RNPs remain enigmatic. In this review we will discuss the function and organization of small RNPs that exist as stable multimers, including RNPs catalyzing RNA chemical modifications, telomerase RNP, and RNPs involved in pre-mRNA splicing.
Conflict of interest statement
Figures

Architecture of box C/D s(no)RNPs. A) Box C/D s(no)RNAs contain conserved sequences, named boxes C, D, C′, and D′. The spacer sequences between boxes C and D′ as well as between boxes C′ and D contain guide sequences that are complementary to the substrate RNAs targeted for methylation. The 5′ and 3′ sequences usually form a terminal stem. B) Conventional mono-sRNP model for archaeal box C/D sRNPs. According to this model, archaeal box C/D sRNPs are composed of one sRNA molecule and two copies of each of the three archaeal box C/D core proteins, assembled symmetrically on the sRNA. C) Di-sRNP model of archaeal box C/D sRNP architecture. According to this structure-based model, archaeal box C/D sRNPs contain two RNA molecules and four sets of each core protein. Colors are: grey - RNA, yellow - L7Ae, blue - Nop5, orange - fibrillarin. D) EM structure of a M. jannaschii box C/D sRNP (EMD-1636, Bleichert et al., 2009) with fitted crystal structures of core proteins (PDB 2nnw, Oruganti et al., 2007; PDB 1xbi, Suryadi et al., 2005). Figures 1A–D are reprinted from (Bleichert et al., 2009). E) and F) Asymmetric assembly models of eukaryotic box C/D mono and di-snoRNP architecture, respectively. Colors are: grey -RNA, yellow - 15.5K/Snu13, light blue - Nop56, dark blue - Nop58, orange - fibrillarin. A color version of this figure is available online.

Telomerase RNP function. A) Domain organization of telomerase reverse transcriptase (TERT). B) Repeat addition cycle for telomerase. Telomerase RNA base pairs to 3′ single stranded overhangs of chromosome ends and TERT reverse transcribes the template sequence into the telomeric DNA. Once the template boundary is reached, telomerase can translocate and catalyze the addition of another telomere repeat, which involves realignment of the 3′ template sequence of telomerase RNA with the newly synthesized repeat sequence. A color version of this figure is available online.

Assembly and recycling of U4/U6.U5 tri-snRNP during pre-mRNA splicing. The U4/U6 di-snRNP is formed by association of the U4 and U6 snRNPs through base pairing of their snRNA components. Subsequently, the U5 snRNP joins to form the U4/U6.U5 tri-snRNP. Recruitment of the U4/U6.U5 tri-snRNPs into the spliceosome results in formation of the B complex. Activation of the spliceosome requires structural rearrangements that result in release of the U4 snRNP and the U6 snRNA associated Lsm2-8 proteins. After splicing is completed, U5 and U6 snRNPs are recycled, joining the U4 snRNP to form U4/U6 di-snRNPs and U4/U6.U5 tri-snRNPs, respectively. A color version of this figure is available online.
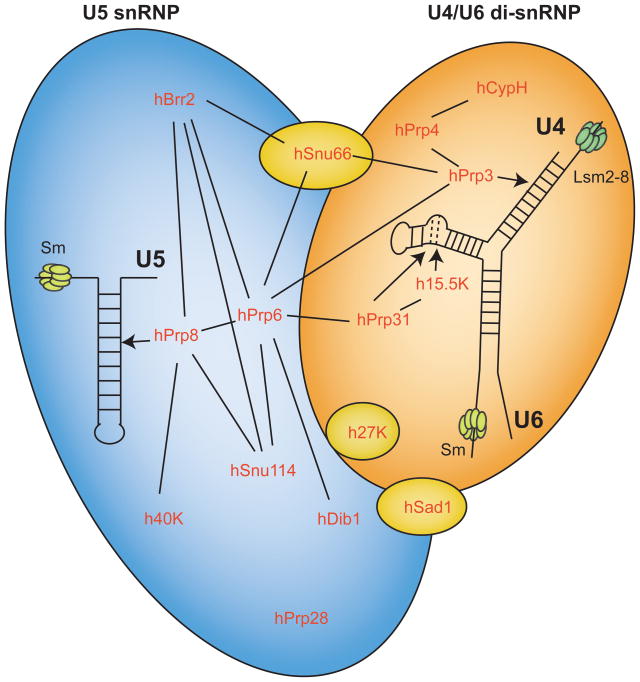
Composition of the human U4/U6.U5 tri-sRNP. U4/U6 di-snRNP (orange) and U5 snRNP (blue) associate to form the U4/U6.U5 tri-snRNP. Proteins specific to the tri-snRNP that are not found in the U4/U6 di-snRNP or U5 snRNP are shown in yellow. Protein-protein interactions within and between RNPs are indicated by solid lines. Known snRNA-protein interactions are indicated by arrows. The U4 and U6 snRNAs are base paired in both the U4/U6 di-snRNP and the U4/U6.U5 tri-sRNP. Proteins are conserved in yeast except for 27K, 40K, and hCypH. A color version of this figure is available online.
Similar articles
-
Independent deposition of heterogeneous nuclear ribonucleoproteins and small nuclear ribonucleoprotein particles at sites of transcription.Proc Natl Acad Sci U S A. 1992 Sep 15;89(18):8409-13. doi: 10.1073/pnas.89.18.8409. Proc Natl Acad Sci U S A. 1992. PMID: 1388268 Free PMC article.
-
Splicing of messenger RNA precursors is inhibited by antisera to small nuclear ribonucleoprotein.Cell. 1983 Nov;35(1):101-7. doi: 10.1016/0092-8674(83)90212-x. Cell. 1983. PMID: 6194895
-
Isolation of distinct small ribonucleoprotein particles containing the spliced leader and U2 RNAs of Trypanosoma brucei.J Biol Chem. 1990 Jun 25;265(18):10582-8. J Biol Chem. 1990. PMID: 2141334
-
Ribonucleoproteins in archaeal pre-rRNA processing and modification.Archaea. 2013;2013:614735. doi: 10.1155/2013/614735. Epub 2013 Mar 10. Archaea. 2013. PMID: 23554567 Free PMC article. Review.
-
Pumping RNA: nuclear bodybuilding along the RNP pipeline.Curr Opin Cell Biol. 2006 Jun;18(3):317-24. doi: 10.1016/j.ceb.2006.03.005. Epub 2006 May 2. Curr Opin Cell Biol. 2006. PMID: 16632338 Review.
Cited by
-
Coarse-Grained Models of RNA Nanotubes for Large Time Scale Studies in Biomedical Applications.Biomedicines. 2020 Jul 6;8(7):195. doi: 10.3390/biomedicines8070195. Biomedicines. 2020. PMID: 32640509 Free PMC article.
-
Systematic Discovery of Endogenous Human Ribonucleoprotein Complexes.Cell Rep. 2019 Oct 29;29(5):1351-1368.e5. doi: 10.1016/j.celrep.2019.09.060. Cell Rep. 2019. PMID: 31665645 Free PMC article.
-
The RNA Revolution in the Central Molecular Biology Dogma Evolution.Int J Mol Sci. 2024 Nov 26;25(23):12695. doi: 10.3390/ijms252312695. Int J Mol Sci. 2024. PMID: 39684407 Free PMC article. Review.
-
MARVEL: an integrated alternative splicing analysis platform for single-cell RNA sequencing data.Nucleic Acids Res. 2023 Mar 21;51(5):e29. doi: 10.1093/nar/gkac1260. Nucleic Acids Res. 2023. PMID: 36631981 Free PMC article.
-
Proteome-Wide Identification of RNA-dependent proteins and an emerging role for RNAs in Plasmodium falciparum protein complexes.Nat Commun. 2024 Feb 14;15(1):1365. doi: 10.1038/s41467-024-45519-1. Nat Commun. 2024. PMID: 38355719 Free PMC article.
References
-
- Achsel T, Ahrens K, Brahms H, Teigelkamp S, Luhrmann R. The human U5-220kD protein (hPrp8) forms a stable RNA-free complex with several U5-specific proteins, including an RNA unwindase, a homologue of ribosomal elongation factor EF-2, and a novel WD-40 protein. Mol Cell Biol. 1998;18:6756–6766. - PMC - PubMed
-
- Aigner S, Postberg J, Lipps HJ, Cech TR. The Euplotes La motif protein p43 has properties of a telomerase-specific subunit. Biochemistry. 2003;42:5736–5747. - PubMed
-
- Aittaleb M, Rashid R, Chen Q, Palmer JR, Daniels CJ, Li H. Structure and function of archaeal box C/D sRNP core proteins. Nat Struct Biol. 2003;10:256–263. - PubMed
-
- Ali MH, Taylor CM, Grigoryan G, Allen KN, Imperiali B, Keating AE. Design of a heterospecific, tetrameric, 21-residue miniprotein with mixed alpha/beta structure. Structure. 2005;13:225–234. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources