

FfVel1 and FfLae1, components of a velvet-like complex in Fusarium fujikuroi, affect differentiation, secondary metabolism and virulence
- PMID: 20572938
- PMCID: PMC2989987
- DOI: 10.1111/j.1365-2958.2010.07263.x
FfVel1 and FfLae1, components of a velvet-like complex in Fusarium fujikuroi, affect differentiation, secondary metabolism and virulence
Abstract
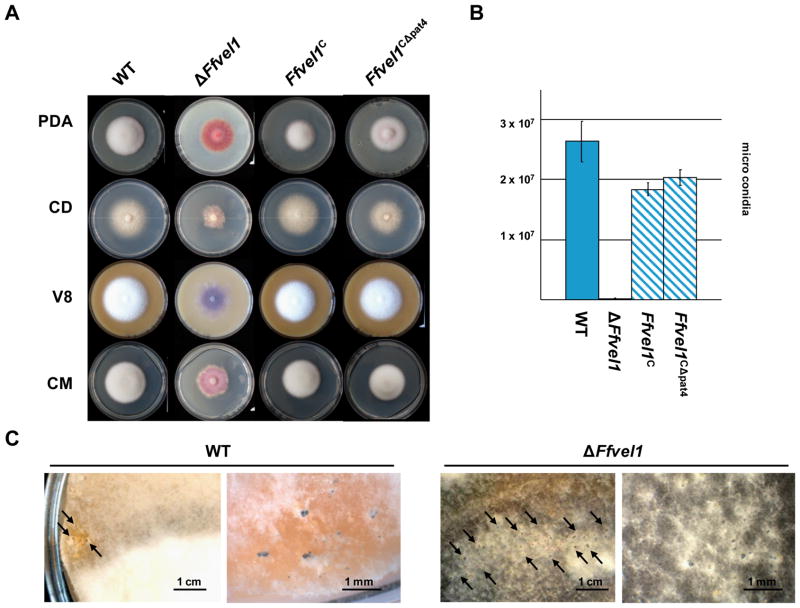
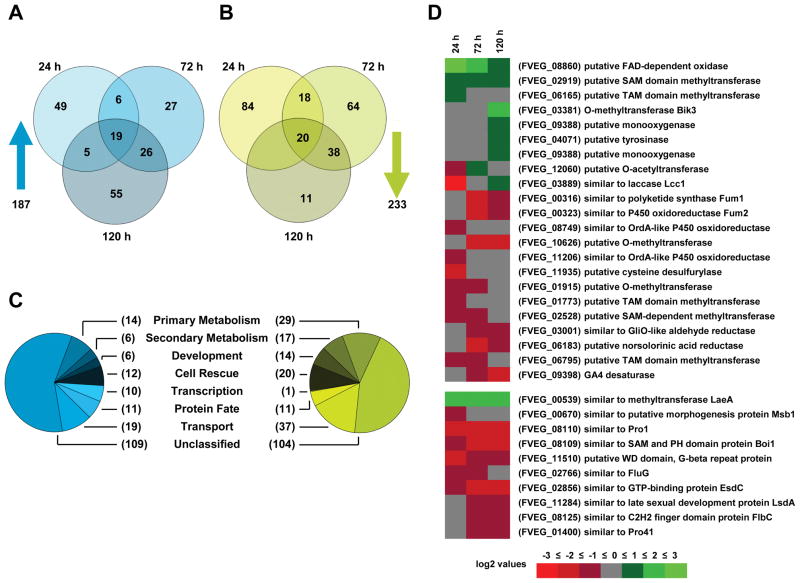
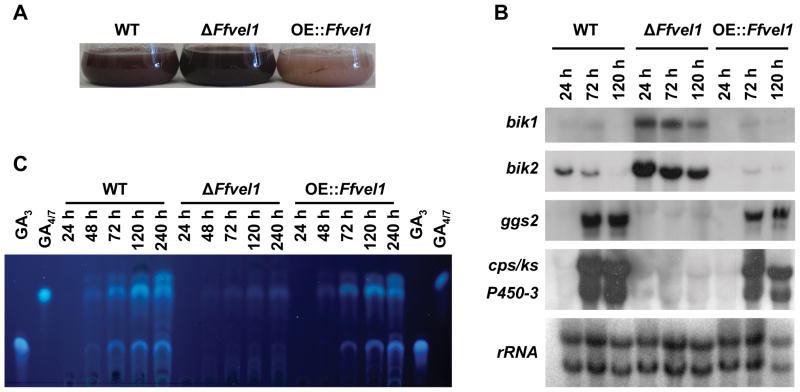
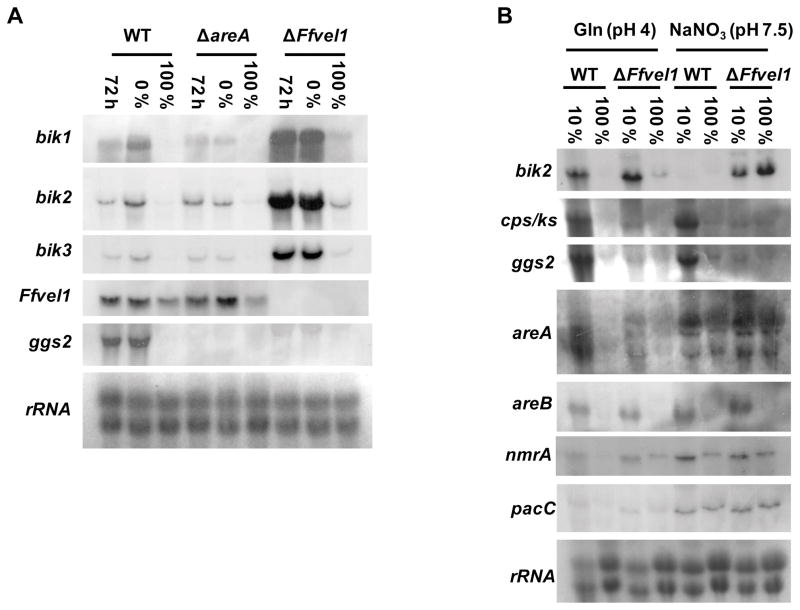
Besides industrially produced gibberellins (GAs), Fusarium fujikuroi is able to produce additional secondary metabolites such as the pigments bikaverin and neurosporaxanthin and the mycotoxins fumonisins and fusarin C. The global regulation of these biosynthetic pathways is only poorly understood. Recently, the velvet complex containing VeA and several other regulatory proteins was shown to be involved in global regulation of secondary metabolism and differentiation in Aspergillus nidulans. Here, we report on the characterization of two components of the F. fujikuroi velvet-like complex, FfVel1 and FfLae1. The gene encoding this first reported LaeA orthologue outside the class of Eurotiomycetidae is upregulated in ΔFfvel1 microarray-studies and FfLae1 interacts with FfVel1 in the nucleus. Deletion of Ffvel1 and Fflae1 revealed for the first time that velvet can simultaneously act as positive (GAs, fumonisins and fusarin C) and negative (bikaverin) regulator of secondary metabolism, and that both components affect conidiation and virulence of F. fujikuroi. Furthermore, the velvet-like protein FfVel2 revealed similar functions regarding conidiation, secondary metabolism and virulence as FfVel1. Cross-genus complementation studies of velvet complex component mutants between Fusarium, Aspergillus and Penicillium support an ancient origin for this complex, which has undergone a divergence in specific functions mediating development and secondary metabolism.
© 2010 Blackwell Publishing Ltd.
Figures

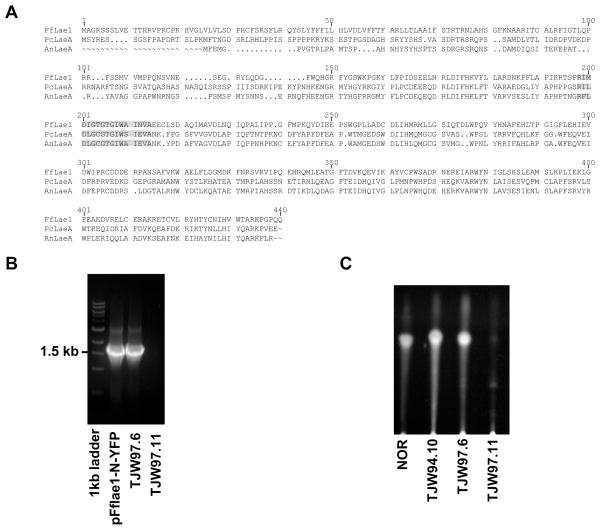
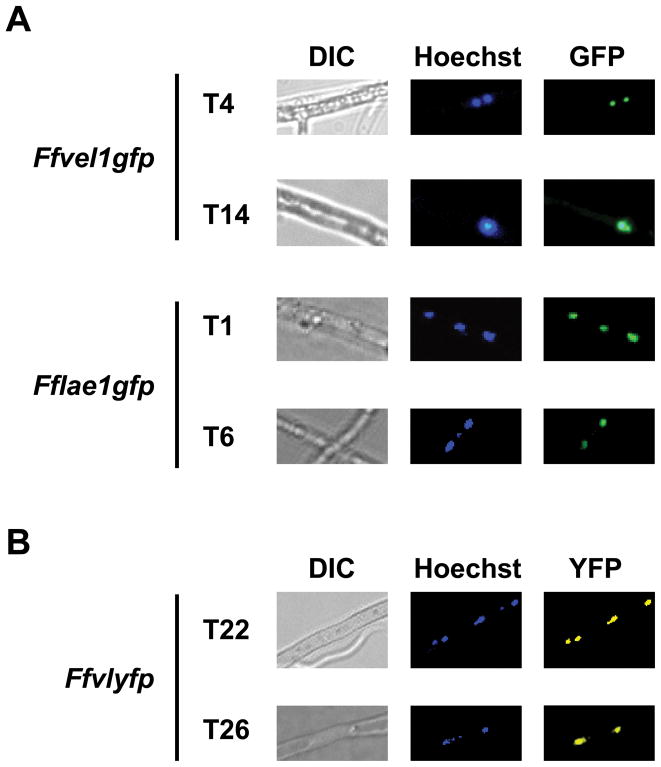
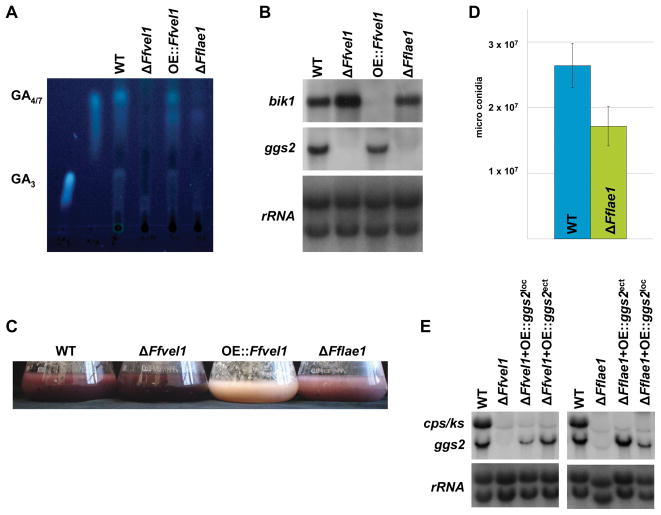
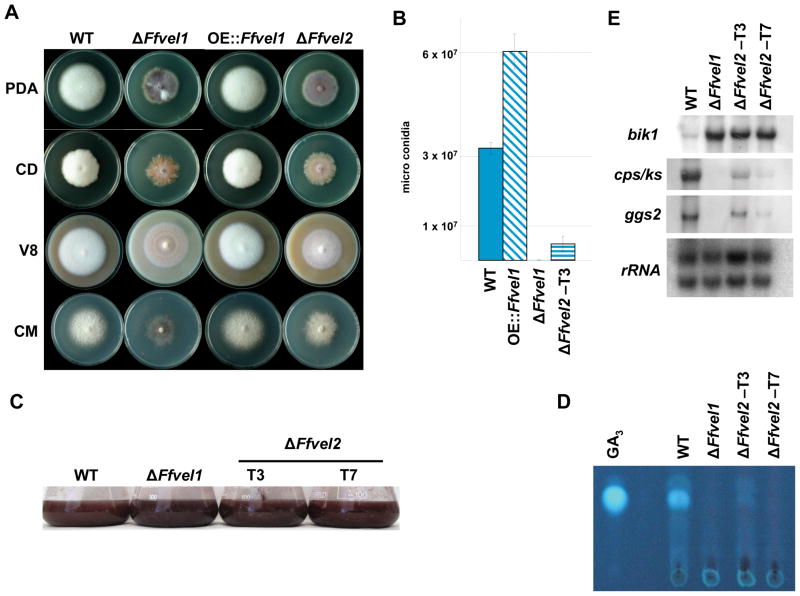
References
-
- Altschul SF, Gish W, Miller W, Myers W, Lipman DJ. Basic local alignment search tool. J Mol Biol. 1990;251:403–410. - PubMed
-
- Ausubel FM, Brent R, Kingston RE, Moore DD, Seidman JG, Smith JA, Struhl K. Current Protocols in Molecular Biology. John Wiley and Sons; New York, NY: 1987.
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
