

Semen-mediated enhancement of HIV infection is donor-dependent and correlates with the levels of SEVI
- PMID: 20573198
- PMCID: PMC2914040
- DOI: 10.1186/1742-4690-7-55
Semen-mediated enhancement of HIV infection is donor-dependent and correlates with the levels of SEVI
Abstract
Background: HIV-1 is usually transmitted in the presence of semen. We have shown that semen boosts HIV-1 infection and contains fragments of prostatic acid phosphatase (PAP) forming amyloid aggregates termed SEVI (semen-derived enhancer of viral infection) that promote virion attachment to target cells. Despite its importance for the global spread of HIV-1, however, the effect of semen on virus infection is controversial.
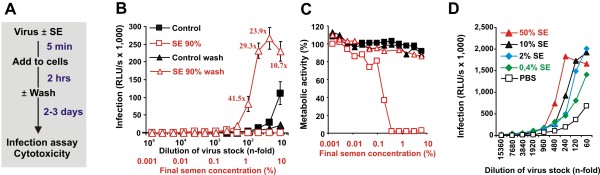
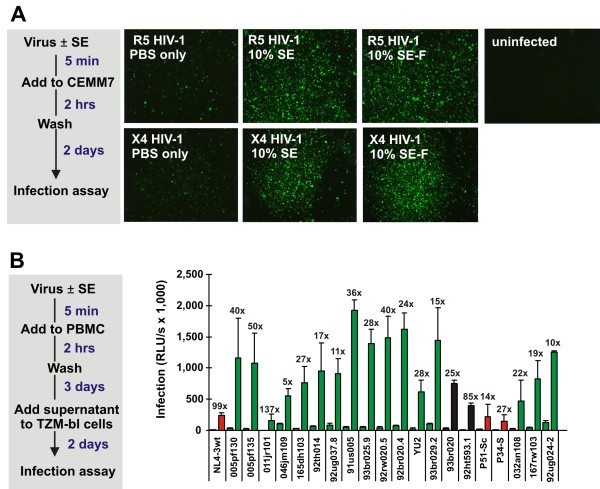
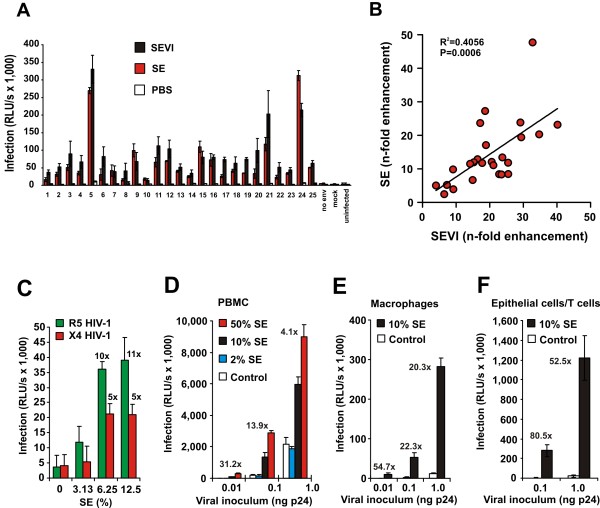
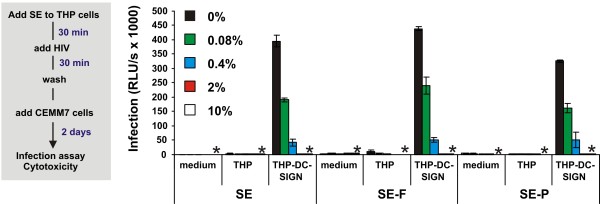
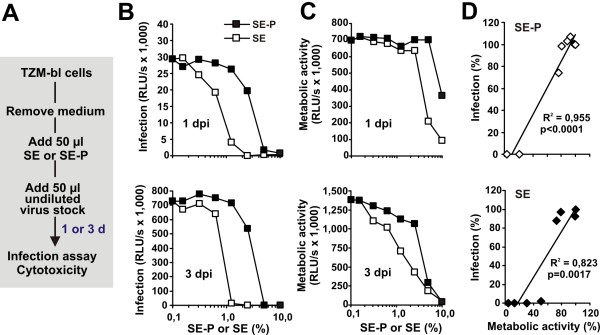
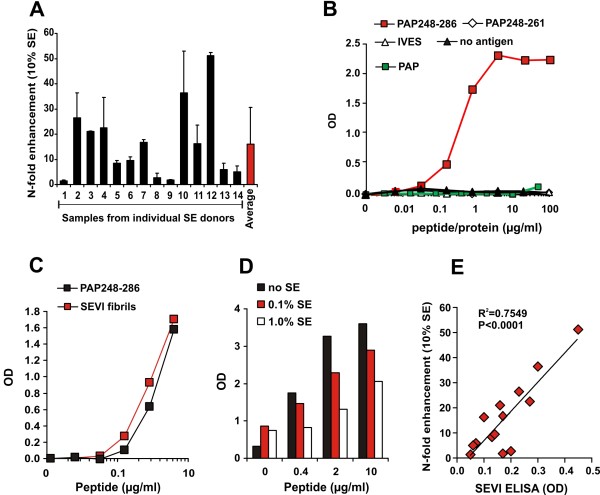
Results: Here, we established methods allowing the meaningful analysis of semen by minimizing its cytotoxic effects and partly recapitulating the conditions encountered during sexual HIV-1 transmission. We show that semen rapidly and effectively enhances the infectivity of HIV-1, HIV-2, and SIV. This enhancement occurs independently of the viral genotype and coreceptor tropism as well as the virus producer and target cell type. Semen-mediated enhancement of HIV-1 infection was also observed under acidic pH conditions and in the presence of vaginal fluid. We further show that the potency of semen in boosting HIV-1 infection is donor dependent and correlates with the levels of SEVI.
Conclusions: Our results show that semen strongly enhances the infectivity of HIV-1 and other primate lentiviruses and that SEVI contributes to this effect. Thus, SEVI may play an important role in the sexual transmission of HIV-1 and addition of SEVI inhibitors to microbicides may improve their efficacy.
Figures

References
-
- UNAIDS. 2008 AIDS epidemic update. http://www.unaids.org
-
- Gray RH, Wawer MJ, Brookmeyer R, Sewankambo NK, Serwadda D, Wabwire-Mangen F, Lutalo T, Li X, vanCott T, Quinn TC. Rakai Project Team. Probability of HIV-1 transmission per coital act in monogamous, heterosexual, HIV-1-discordant couples in Rakai, Uganda. Lancet. 2001;357:1149–53. doi: 10.1016/S0140-6736(00)04331-2. - DOI - PubMed
-
- Padian NS, Shiboski SC, Glass SO, Vittinghoff E. Heterosexual transmission of human immunodeficiency virus (HIV) in northern California: results from a ten-year study. Am J Epidemiol. 1997;146:350–57. - PubMed
-
- Vittinghoff E, Douglas J, Judson F, McKirnan D, MacQueen K, Buchbinder S. Per-contact risk of human immunodeficiency virus transmission between male sexual partners. Am J Epidemiol. 1999;150:306–11. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
