

Alternative splicing is frequent during early embryonic development in mouse
- PMID: 20573213
- PMCID: PMC2898759
- DOI: 10.1186/1471-2164-11-399
Alternative splicing is frequent during early embryonic development in mouse
Abstract
Background: Alternative splicing is known to increase the complexity of mammalian transcriptomes since nearly all mammalian genes express multiple pre-mRNA isoforms. However, our knowledge of the extent and function of alternative splicing in early embryonic development is based mainly on a few isolated examples. High throughput technologies now allow us to study genome-wide alternative splicing during mouse development.
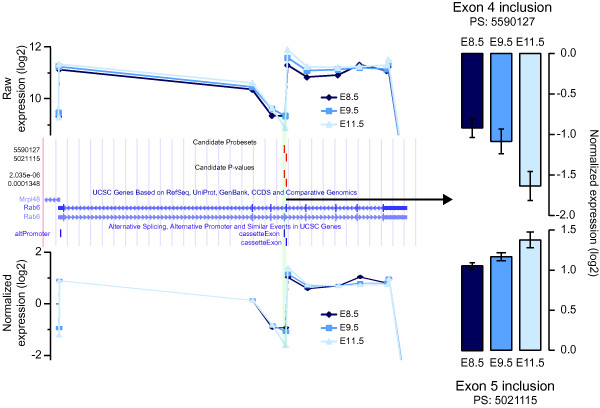
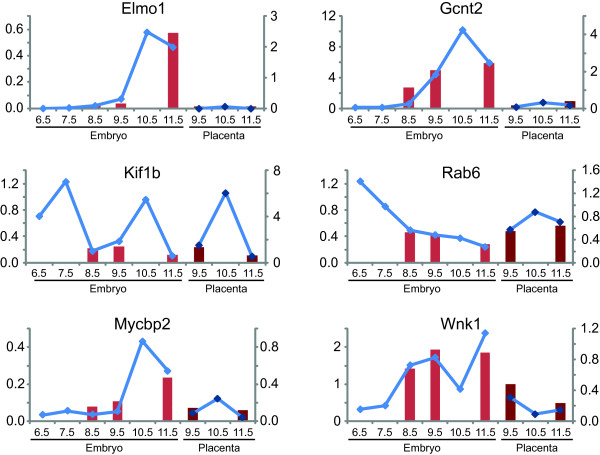
Results: A genome-wide analysis of alternative isoform expression in embryonic day 8.5, 9.5 and 11.5 mouse embryos and placenta was carried out using a splicing-sensitive exon microarray. We show that alternative splicing and isoform expression is frequent across developmental stages and tissues, and is comparable in frequency to the variation in whole-transcript expression. The genes that are alternatively spliced across our samples are disproportionately involved in important developmental processes. Finally, we find that a number of RNA binding proteins, including putative splicing factors, are differentially expressed and spliced across our samples suggesting that such proteins may be involved in regulating tissue and temporal variation in isoform expression. Using an example of a well characterized splicing factor, Fox2, we demonstrate that changes in Fox2 expression levels can be used to predict changes in inclusion levels of alternative exons that are flanked by Fox2 binding sites.
Conclusions: We propose that alternative splicing is an important developmental regulatory mechanism. We further propose that gene expression should routinely be monitored at both the whole transcript and the isoform level in developmental studies.
Figures
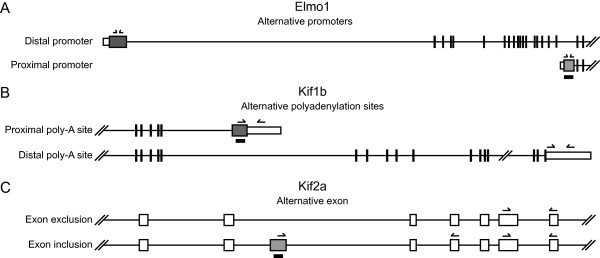
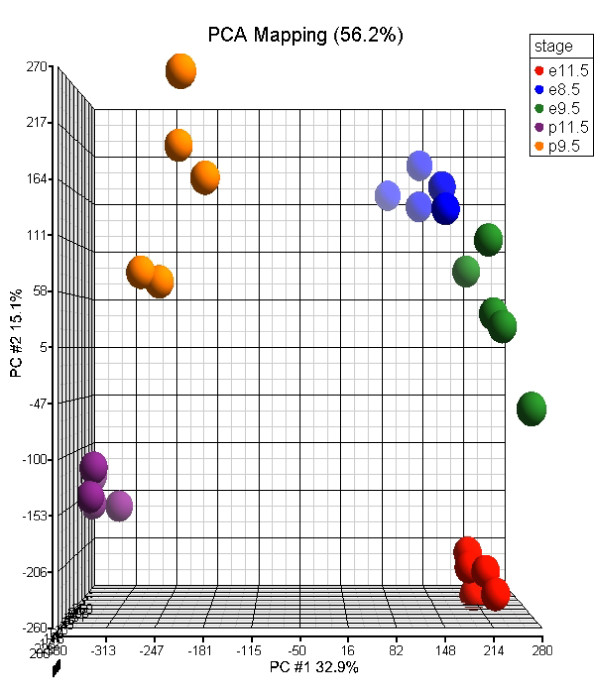
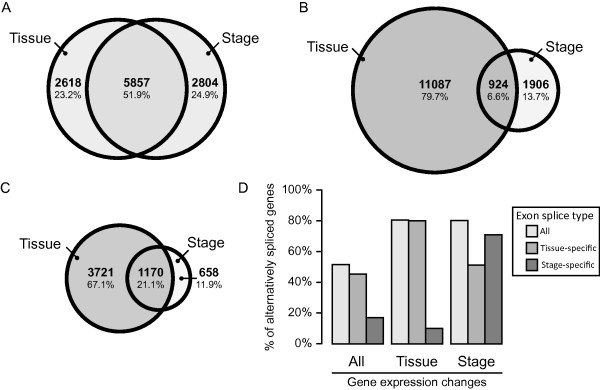
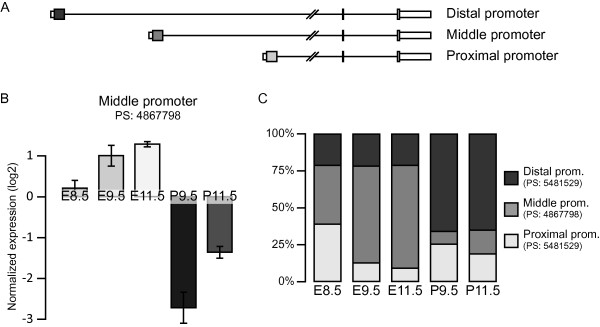

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
