

Distinct molecular pathways to X4 tropism for a V3-truncated human immunodeficiency virus type 1 lead to differential coreceptor interactions and sensitivity to a CXCR4 antagonist
- PMID: 20573813
- PMCID: PMC2919036
- DOI: 10.1128/JVI.00333-10
Distinct molecular pathways to X4 tropism for a V3-truncated human immunodeficiency virus type 1 lead to differential coreceptor interactions and sensitivity to a CXCR4 antagonist
Abstract
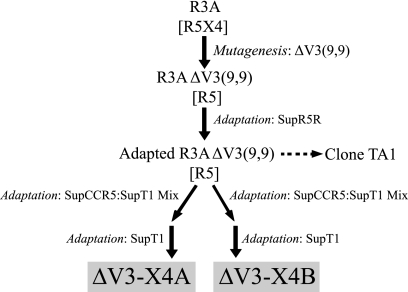
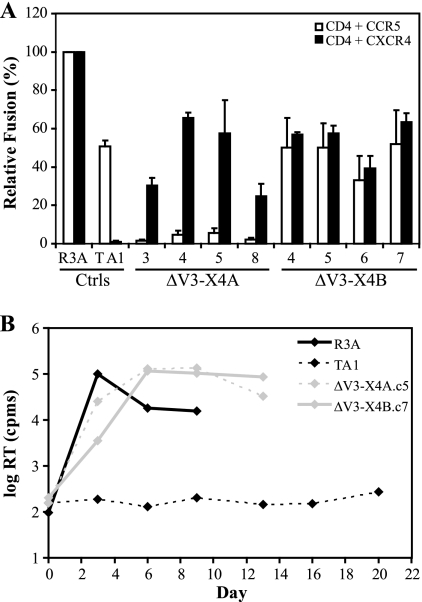
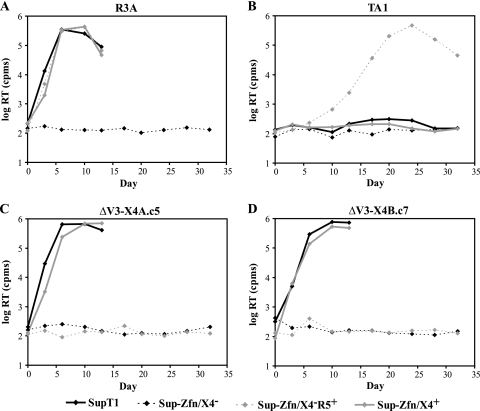
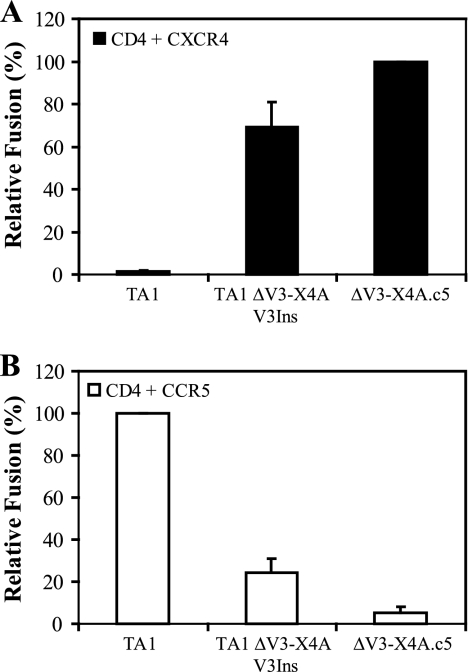
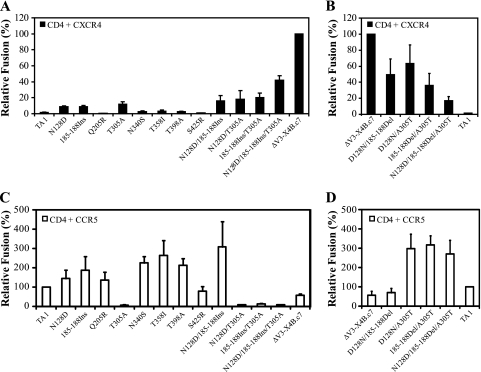
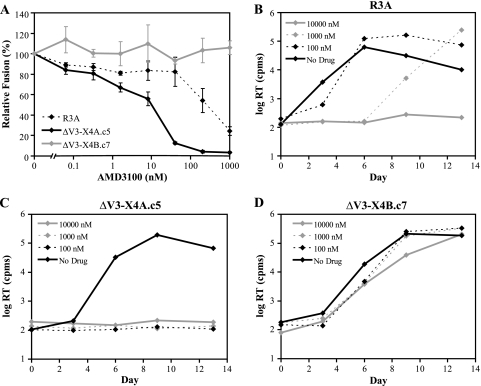
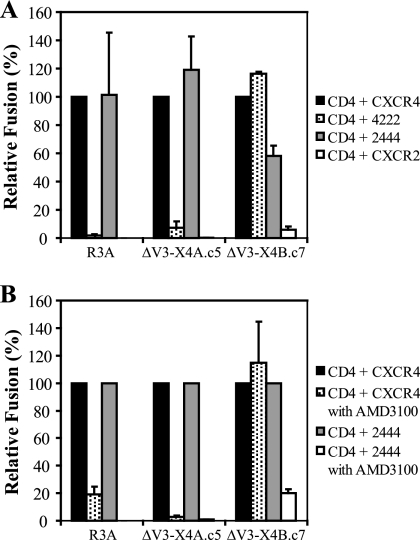
During the course of infection, transmitted HIV-1 isolates that initially use CCR5 can acquire the ability to use CXCR4, which is associated with an accelerated progression to AIDS. Although this coreceptor switch is often associated with mutations in the stem of the viral envelope (Env) V3 loop, domains outside V3 can also play a role, and the underlying mechanisms and structural basis for how X4 tropism is acquired remain unknown. In this study we used a V3 truncated R5-tropic Env as a starting point to derive two X4-tropic Envs, termed DeltaV3-X4A.c5 and DeltaV3-X4B.c7, which took distinct molecular pathways for this change. The DeltaV3-X4A.c5 Env clone acquired a 7-amino-acid insertion in V3 that included three positively charged residues, reestablishing an interaction with the CXCR4 extracellular loops (ECLs) and rendering it highly susceptible to the CXCR4 antagonist AMD3100. In contrast, the DeltaV3-X4B.c7 Env maintained the V3 truncation but acquired mutations outside V3 that were critical for X4 tropism. In contrast to DeltaV3-X4A.c5, DeltaV3-X4B.c7 showed increased dependence on the CXCR4 N terminus (NT) and was completely resistant to AMD3100. These results indicate that HIV-1 X4 coreceptor switching can involve (i) V3 loop mutations that establish interactions with the CXCR4 ECLs, and/or (ii) mutations outside V3 that enhance interactions with the CXCR4 NT. The cooperative contributions of CXCR4 NT and ECL interactions with gp120 in acquiring X4 tropism likely impart flexibility on pathways for viral evolution and suggest novel approaches to isolate these interactions for drug discovery.
Figures

Similar articles
-
Effects of partial deletions within the human immunodeficiency virus type 1 V3 loop on coreceptor tropism and sensitivity to entry inhibitors.J Virol. 2008 Jan;82(2):664-73. doi: 10.1128/JVI.01793-07. Epub 2007 Oct 31. J Virol. 2008. PMID: 17977968 Free PMC article.
-
Tissue-specific sequence alterations in the human immunodeficiency virus type 1 envelope favoring CCR5 usage contribute to persistence of dual-tropic virus in the brain.J Virol. 2009 Jun;83(11):5430-41. doi: 10.1128/JVI.02648-08. Epub 2009 Mar 25. J Virol. 2009. PMID: 19321618 Free PMC article.
-
Differences in molecular evolution between switch (R5 to R5X4/X4-tropic) and non-switch (R5-tropic only) HIV-1 populations during infection.Infect Genet Evol. 2010 Apr;10(3):356-64. doi: 10.1016/j.meegid.2009.05.003. Epub 2009 May 14. Infect Genet Evol. 2010. PMID: 19446658
-
HIV-1 subtype C predicted co-receptor tropism in Africa: an individual sequence level meta-analysis.AIDS Res Ther. 2020 Feb 7;17(1):5. doi: 10.1186/s12981-020-0263-x. AIDS Res Ther. 2020. PMID: 32033571 Free PMC article. Review.
-
Genotypic coreceptor analysis.Eur J Med Res. 2007 Oct 15;12(9):453-62. Eur J Med Res. 2007. PMID: 17933727 Review.
Cited by
-
Study of compartmentalization in the visna clinical form of small ruminant lentivirus infection in sheep.BMC Vet Res. 2012 Jan 26;8:8. doi: 10.1186/1746-6148-8-8. BMC Vet Res. 2012. PMID: 22281181 Free PMC article.
-
A Highly Unusual V1 Region of Env in an Elite Controller of HIV Infection.J Virol. 2019 May 1;93(10):e00094-19. doi: 10.1128/JVI.00094-19. Print 2019 May 15. J Virol. 2019. PMID: 30842322 Free PMC article.
-
Potent and Broad Inhibition of HIV-1 by a Peptide from the gp41 Heptad Repeat-2 Domain Conjugated to the CXCR4 Amino Terminus.PLoS Pathog. 2016 Nov 17;12(11):e1005983. doi: 10.1371/journal.ppat.1005983. eCollection 2016 Nov. PLoS Pathog. 2016. PMID: 27855210 Free PMC article.
-
Stochastic model of in-vivo X4 emergence during HIV infection: implications for the CCR5 inhibitor maraviroc.PLoS One. 2012;7(7):e38755. doi: 10.1371/journal.pone.0038755. Epub 2012 Jul 17. PLoS One. 2012. PMID: 22866173 Free PMC article.
-
Interaction of the HIV-1 gp120 viral protein V3 loop with bacterial lipopolysaccharide: a pattern recognition inhibition.J Biol Chem. 2011 Jul 22;286(29):26228-37. doi: 10.1074/jbc.M111.220434. Epub 2011 Jun 2. J Biol Chem. 2011. PMID: 21636577 Free PMC article.
References
-
- Agrawal-Gamse, C., F. H. Lee, B. Haggarty, A. P. Jordan, Y. Yi, B. Lee, R. G. Collman, J. A. Hoxie, R. W. Doms, and M. M. Laakso. 2009. Adaptive mutations in a human immunodeficiency virus type 1 envelope protein with a truncated V3 loop restore function by improving interactions with CD4. J. Virol. 83:11005-11015. - PMC - PubMed
-
- Alkhatib, G., C. Combadiere, C. C. Broder, Y. Feng, P. E. Kennedy, P. M. Murphy, and E. A. Berger. 1996. CC CKR5: a RANTES, MIP-1α, MIP-1β receptor as a fusion cofactor for macrophage-tropic HIV-1. Science 272:1955-1958. - PubMed
-
- Atchison, R. E., J. Gosling, F. S. Monteclaro, C. Franci, L. Digilio, I. F. Charo, and M. A. Goldsmith. 1996. Multiple extracellular elements of CCR5 and HIV-1 entry: dissociation from response to chemokines. Science 274:1924-1926. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
