

A single asparagine-linked glycosylation site of the severe acute respiratory syndrome coronavirus spike glycoprotein facilitates inhibition by mannose-binding lectin through multiple mechanisms
- PMID: 20573835
- PMCID: PMC2919028
- DOI: 10.1128/JVI.00554-10
A single asparagine-linked glycosylation site of the severe acute respiratory syndrome coronavirus spike glycoprotein facilitates inhibition by mannose-binding lectin through multiple mechanisms
Abstract
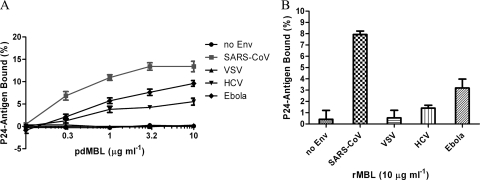
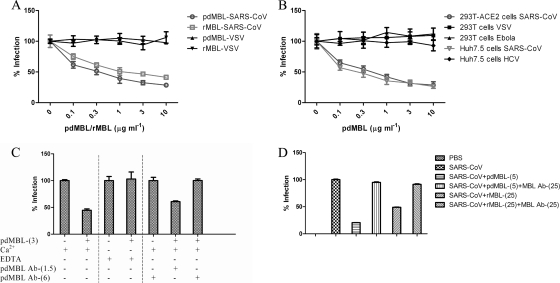
Mannose-binding lectin (MBL) is a serum protein that plays an important role in host defenses as an opsonin and through activation of the complement system. The objective of this study was to assess the interactions between MBL and severe acute respiratory syndrome-coronavirus (SARS-CoV) spike (S) glycoprotein (SARS-S). MBL was found to selectively bind to retroviral particles pseudotyped with SARS-S. Unlike several other viral envelopes to which MBL can bind, both recombinant and plasma-derived human MBL directly inhibited SARS-S-mediated viral infection. Moreover, the interaction between MBL and SARS-S blocked viral binding to the C-type lectin, DC-SIGN. Mutagenesis indicated that a single N-linked glycosylation site, N330, was critical for the specific interactions between MBL and SARS-S. Despite the proximity of N330 to the receptor-binding motif of SARS-S, MBL did not affect interactions with the ACE2 receptor or cathepsin L-mediated activation of SARS-S-driven membrane fusion. Thus, binding of MBL to SARS-S may interfere with other early pre- or postreceptor-binding events necessary for efficient viral entry.
Figures
References
-
- Beebe, D. P., and N. R. Cooper. 1981. Neutralization of vesicular stomatitis virus (VSV) by human complement requires a natural IgM antibody present in human serum. J. Immunol. 126:1562-1568. - PubMed
-
- Brown, K. S., S. D. Ryder, W. L. Irving, R. B. Sim, and T. P. Hickling. 2007. Mannan binding lectin and viral hepatitis. Immunol. Lett. 108:34-44. - PubMed
-
- Chaipan, C., E. J. Soilleux, P. Simpson, H. Hofmann, T. Gramberg, A. Marzi, M. Geier, E. A. Stewart, J. Eisemann, A. Steinkasserer, K. Suzuki-Inoue, G. L. Fuller, A. C. Pearce, S. P. Watson, J. A. Hoxie, F. Baribaud, and S. Pohlmann. 2006. DC-SIGN and CLEC-2 mediate human immunodeficiency virus type 1 capture by platelets. J. Virol. 80:8951-8960. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
