

Dipeptidyl peptidase-like protein 6 is required for normal electrophysiological properties of cerebellar granule cells
- PMID: 20573902
- PMCID: PMC2916862
- DOI: 10.1523/JNEUROSCI.5489-09.2010
Dipeptidyl peptidase-like protein 6 is required for normal electrophysiological properties of cerebellar granule cells
Abstract
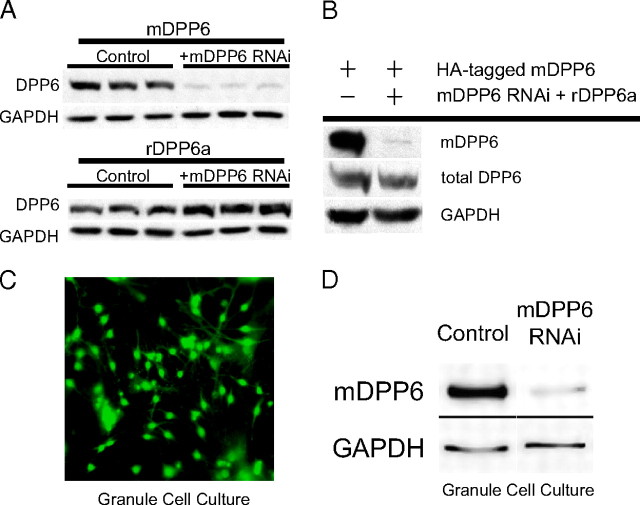
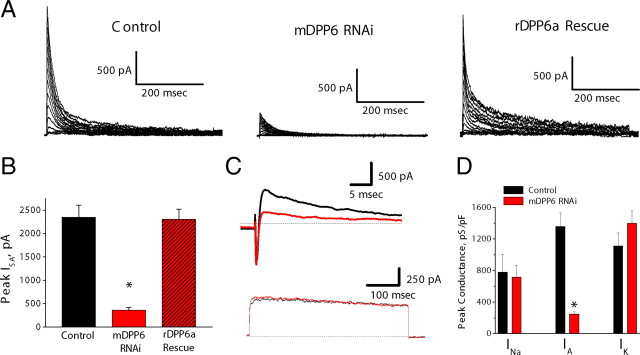
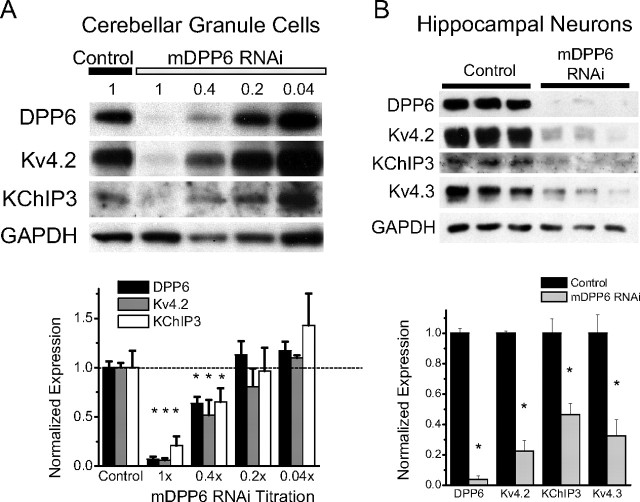
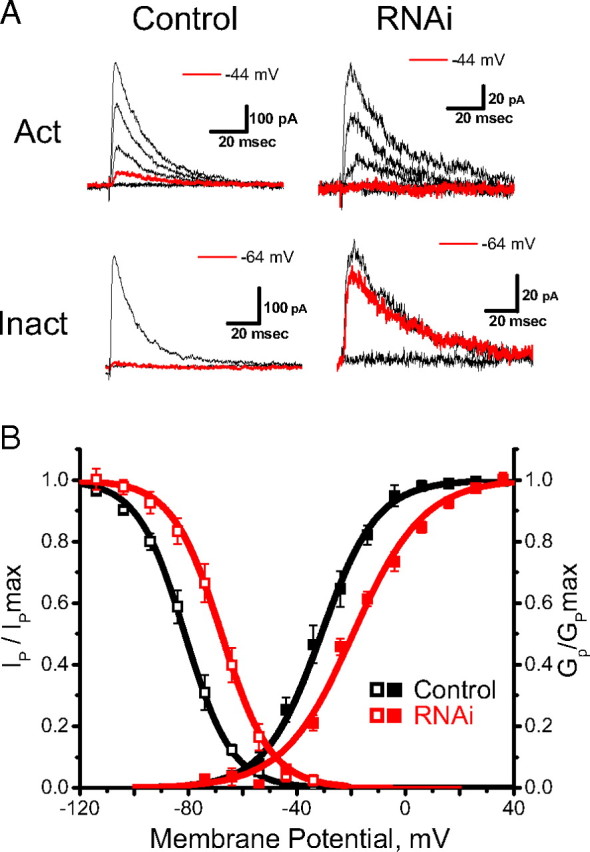
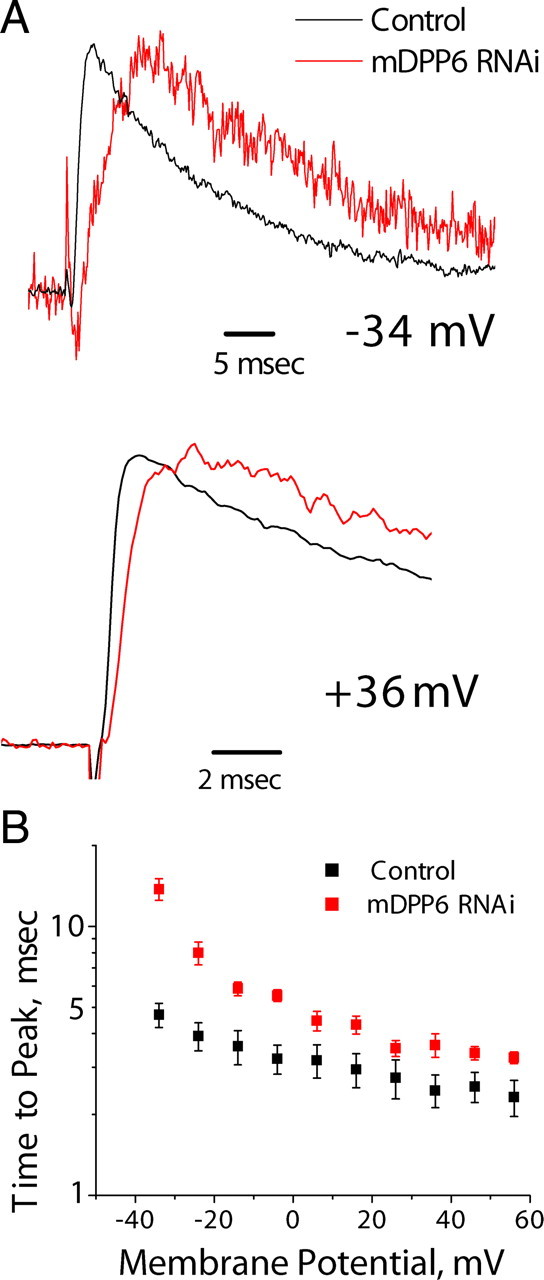
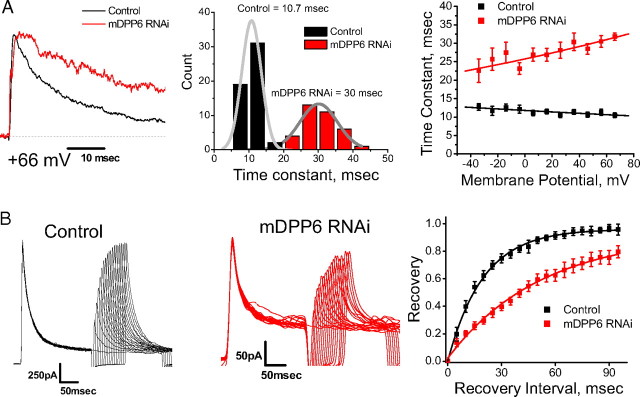
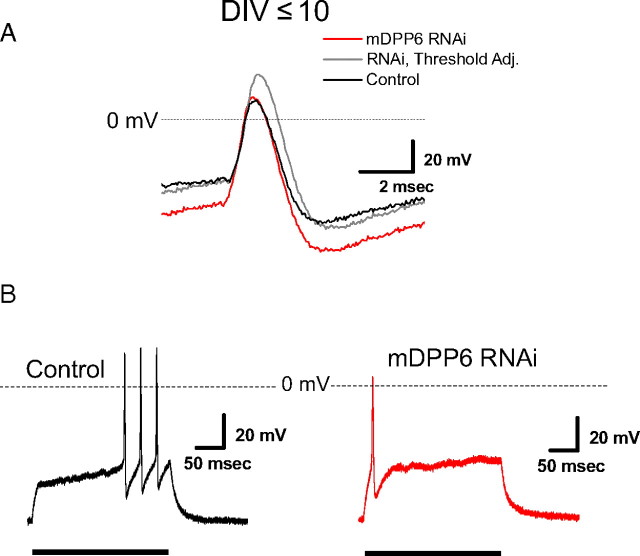
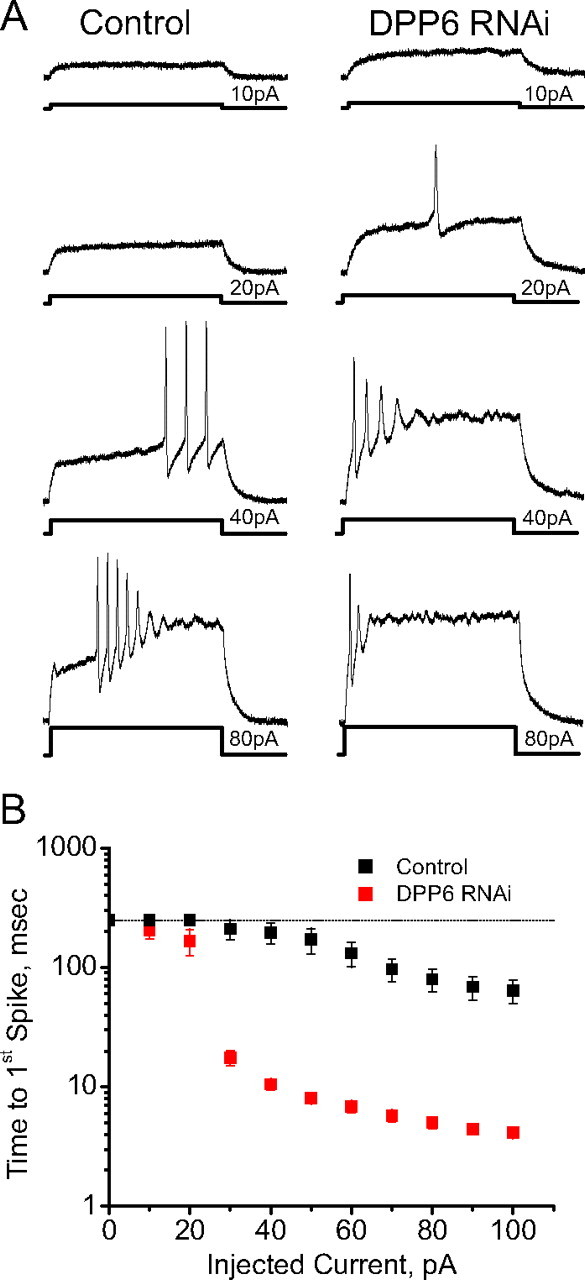
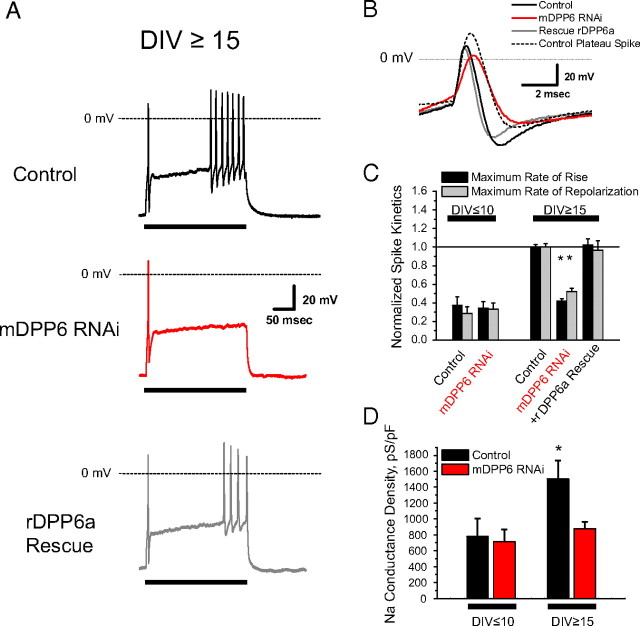
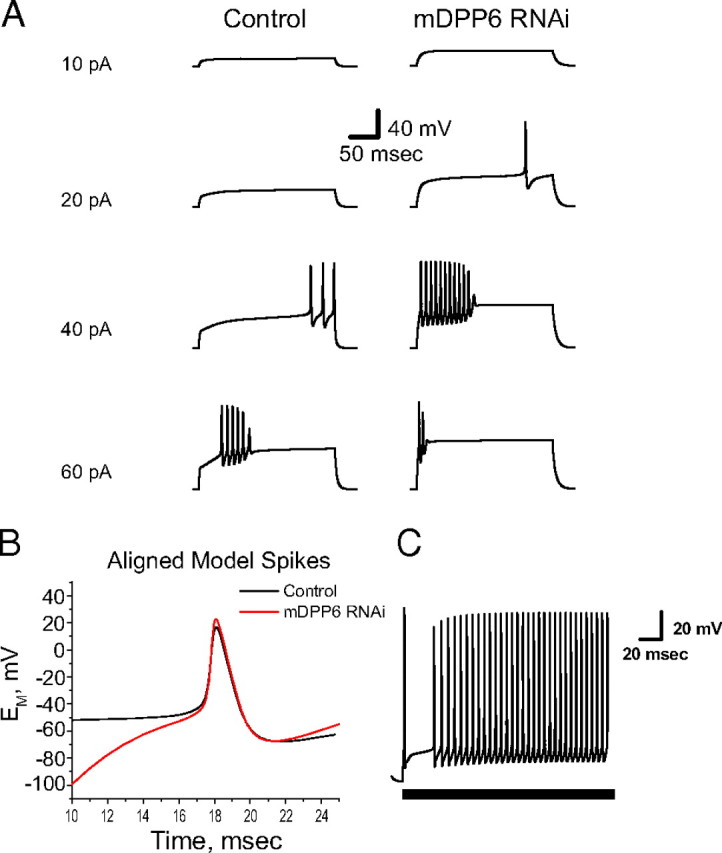
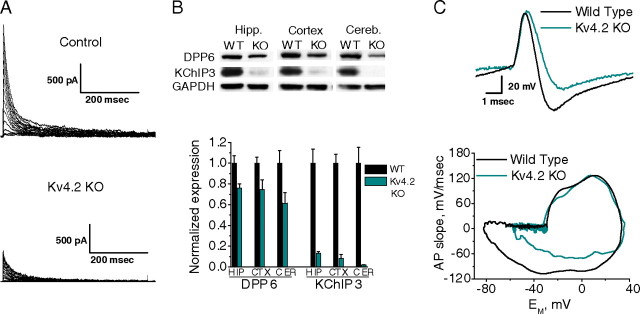
In cerebellar granule (CG) cells and many other neurons, A-type potassium currents play an important role in regulating neuronal excitability, firing patterns, and activity-dependent plasticity. Protein biochemistry has identified dipeptidyl peptidase-like protein 6 (DPP6) as an auxiliary subunit of Kv4-based A-type channels and thus a potentially important regulator of neuronal excitability. In this study, we used an RNA interference (RNAi) strategy to examine the role DPP6 plays in forming and shaping the electrophysiological properties of CG cells. DPP6 RNAi delivered by lentiviral vectors effectively disrupts DPP6 protein expression in CG cells. In response to the loss of DPP6, I(SA) peak conductance amplitude is reduced by >85% in parallel with a dramatic reduction in the level of I(SA) channel protein complex found in CG cells. The I(SA) channels remaining in CG cells after suppression of DPP6 show alterations in gating similar to Kv4 channels expressed in heterologous systems without DPP6. In addition to these effects on A-type current, we find that loss of DPP6 has additional effects on input resistance and Na(+) channel conductance that combine with the effects on I(SA) to produce a global change in excitability. Overall, DPP6 expression seems to be critical for the expression of a high-frequency electrophysiological phenotype in CG cells by increasing leak conductance, A-type current levels and kinetics, and Na(+) current amplitude.
Figures
References
-
- Aamodt SM, Constantine-Paton M. The role of neural activity in synaptic development and its implications for adult brain function. Adv Neurol. 1999;79:133–144. - PubMed
-
- Alders M, Koopmann TT, Christiaans I, Postema PG, Beekman L, Tanck MW, Zeppenfeld K, Loh P, Koch KT, Demolombe S, Mannens MM, Bezzina CR, Wilde AA. Haplotype-sharing analysis implicates chromosome 7q36 harboring DPP6 in familial idiopathic ventricular fibrillation. Am J Hum Genet. 2009;84:468–476. - PMC - PubMed
-
- Allen M, Heinzmann A, Noguchi E, Abecasis G, Broxholme J, Ponting CP, Bhattacharyya S, Tinsley J, Zhang Y, Holt R, Jones EY, Lench N, Carey A, Jones H, Dickens NJ, Dimon C, Nicholls R, Baker C, Xue L, Townsend E, et al. Positional cloning of a novel gene influencing asthma from chromosome 2q14. Nat Genet. 2003;35:258–263. - PubMed
-
- An WF, Bowlby MR, Betty M, Cao J, Ling HP, Mendoza G, Hinson JW, Mattsson KI, Strassle BW, Trimmer JS, Rhodes KJ. Modulation of A-type potassium channels by a family of calcium sensors. Nature. 2000;403:553–556. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases