

Sequential counteracting kinases restrict an asymmetric gene expression program to early G1
- PMID: 20573982
- PMCID: PMC2921117
- DOI: 10.1091/mbc.E10-02-0174
Sequential counteracting kinases restrict an asymmetric gene expression program to early G1
Abstract
Gene expression is restricted to specific times in cell division and differentiation through close control of both activation and inactivation of transcription. In budding yeast, strict spatiotemporal regulation of the transcription factor Ace2 ensures that it acts only once in a cell's lifetime: at the M-to-G1 transition in newborn daughter cells. The Ndr/LATS family kinase Cbk1, functioning in a system similar to metazoan hippo signaling pathways, activates Ace2 and drives its accumulation in daughter cell nuclei, but the mechanism of this transcription factor's inactivation is unknown. We found that Ace2's nuclear localization is maintained by continuous Cbk1 activity and that inhibition of the kinase leads to immediate loss of phosphorylation and export to the cytoplasm. Once exported, Ace2 cannot re-enter nuclei for the remainder of the cell cycle. Two separate mechanisms enforce Ace2's cytoplasmic sequestration: 1) phosphorylation of CDK consensus sites in Ace2 by the G1 CDKs Pho85 and Cdc28/CDK1 and 2) an unknown mechanism mediated by Pho85 that is independent of its kinase activity. Direct phosphorylation of CDK consensus sites is not necessary for Ace2's cytoplasmic retention, indicating that these mechanisms function redundantly. Overall, these findings show how sequential opposing kinases limit a daughter cell specific transcriptional program to a brief period during the cell cycle and suggest that CDKs may function as cytoplasmic sequestration factors.
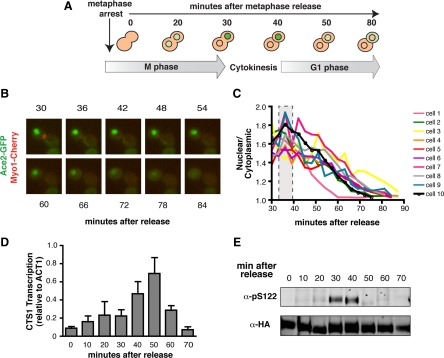
Figures





Similar articles
-
Mitotic exit control of the Saccharomyces cerevisiae Ndr/LATS kinase Cbk1 regulates daughter cell separation after cytokinesis.Mol Cell Biol. 2011 Feb;31(4):721-35. doi: 10.1128/MCB.00403-10. Epub 2010 Dec 6. Mol Cell Biol. 2011. PMID: 21135117 Free PMC article.
-
The NDR/LATS family kinase Cbk1 directly controls transcriptional asymmetry.PLoS Biol. 2008 Aug 19;6(8):e203. doi: 10.1371/journal.pbio.0060203. PLoS Biol. 2008. PMID: 18715118 Free PMC article.
-
The Ndr/LATS Kinase Cbk1 Regulates a Specific Subset of Ace2 Functions and Suppresses the Hypha-to-Yeast Transition in Candida albicans.mBio. 2020 Aug 18;11(4):e01900-20. doi: 10.1128/mBio.01900-20. mBio. 2020. PMID: 32817109 Free PMC article.
-
Cyclin-dependent kinases and S phase control in mammalian cells.Cell Cycle. 2003 Jul-Aug;2(4):316-24. Cell Cycle. 2003. PMID: 12851482 Review.
-
Regulation of the initiation step of DNA replication by cyclin-dependent kinases.Chromosoma. 2010 Dec;119(6):565-74. doi: 10.1007/s00412-010-0291-8. Epub 2010 Aug 5. Chromosoma. 2010. PMID: 20686781 Review.
Cited by
-
Nuclear autonomy in multinucleate fungi.Curr Opin Microbiol. 2015 Dec;28:60-5. doi: 10.1016/j.mib.2015.08.009. Epub 2015 Sep 14. Curr Opin Microbiol. 2015. PMID: 26379197 Free PMC article. Review.
-
Nuclear envelope morphology constrains diffusion and promotes asymmetric protein segregation in closed mitosis.J Cell Biol. 2012 Jun 25;197(7):921-37. doi: 10.1083/jcb.201112117. Epub 2012 Jun 18. J Cell Biol. 2012. PMID: 22711697 Free PMC article.
-
Topology and control of the cell-cycle-regulated transcriptional circuitry.Genetics. 2014 Jan;196(1):65-90. doi: 10.1534/genetics.113.152595. Genetics. 2014. PMID: 24395825 Free PMC article. Review.
-
Mitotic exit control of the Saccharomyces cerevisiae Ndr/LATS kinase Cbk1 regulates daughter cell separation after cytokinesis.Mol Cell Biol. 2011 Feb;31(4):721-35. doi: 10.1128/MCB.00403-10. Epub 2010 Dec 6. Mol Cell Biol. 2011. PMID: 21135117 Free PMC article.
-
The Cbk1-Ace2 axis guides Candida albicans from yeast to hyphae and back again.Curr Genet. 2021 Jun;67(3):461-469. doi: 10.1007/s00294-020-01152-1. Epub 2021 Jan 12. Curr Genet. 2021. PMID: 33433733 Free PMC article.
References
-
- Archambault V., Chang E. J., Drapkin B. J., Cross F. R., Chait B. T., Rout M. P. Targeted proteomic study of the cyclin-cdk module. Mol. Cell. 2004;14:699–711. - PubMed
-
- Artavanis-Tsakonas S., Matsuno K., Fortini M. E. Notch signaling. Science. 1995;268:225–232. - PubMed
-
- Bishop A. C., Buzko O., Shokat K. M. Magic bullets for protein kinases. Trends Cell Biol. 2001;11:167–172. - PubMed
-
- Bishop A. C., Shah K., Liu Y., Witucki L., Kung C., Shokat K. M. Design of allele-specific inhibitors to probe protein kinase signaling. Curr. Biol. 1998;8:257–266. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
