

The {omega}-3 fatty acid eicosapentaenoic acid elicits cAMP generation in colonic epithelial cells via a "store-operated" mechanism
- PMID: 20576916
- PMCID: PMC2950681
- DOI: 10.1152/ajpgi.00028.2010
The {omega}-3 fatty acid eicosapentaenoic acid elicits cAMP generation in colonic epithelial cells via a "store-operated" mechanism
Abstract
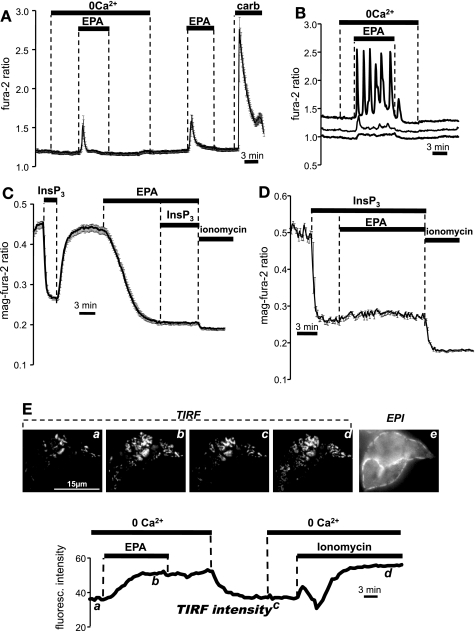
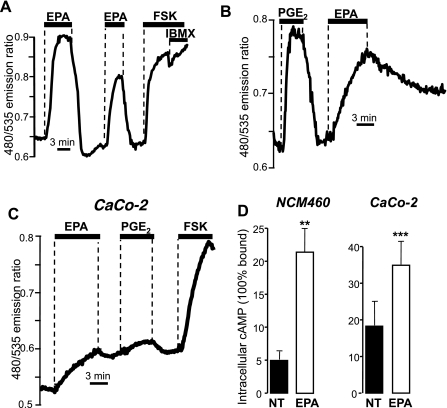
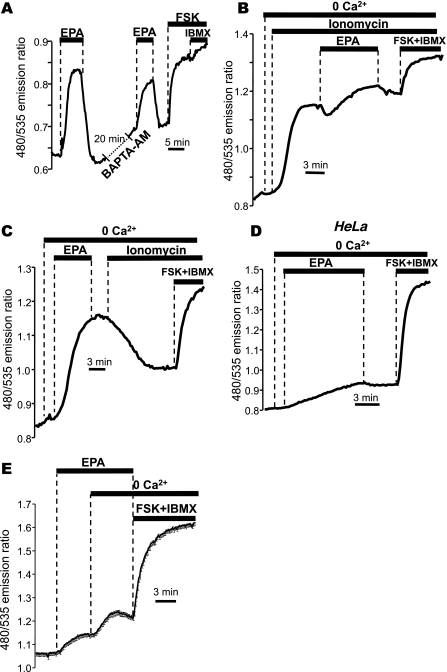
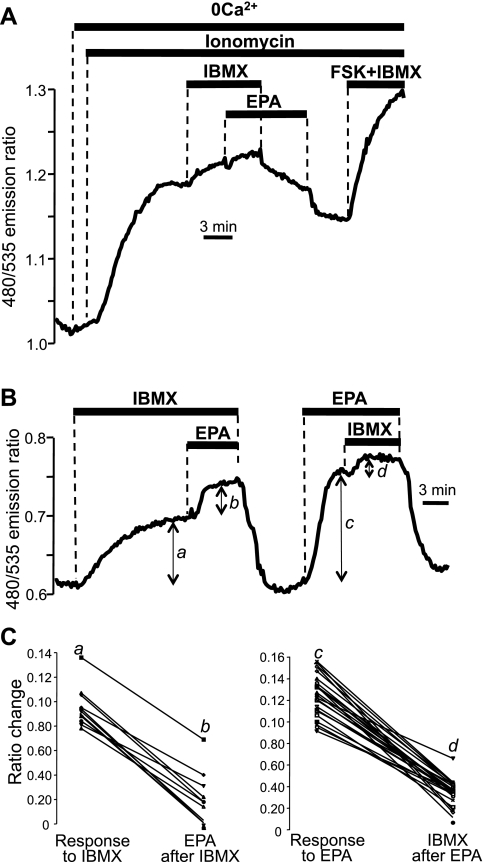
Eicosapentaenoic acid (EPA) is an omega-3 polyunsaturated fatty acid abundant in fish oil that exerts a wide spectrum of documented beneficial health effects in humans. Because dietary interventions are relatively inexpensive and are widely assumed to be safe, they have broad public appeal. Their endorsement can potentially have a major impact on human health, but hard mechanistic evidence that specifies how these derivatives work at the cellular level is limited. EPA (50 microM) caused a small elevation of cytoplasmic Ca(2+) concentration ([Ca(2+)]) in intact NCM460 human colonic epithelial cells as measured by fura 2 and a profound drop of [Ca(2+)] within the endoplasmic reticulum (ER) of permeabilized cells as monitored by compartmentalized mag-fura 2. Total internal reflection fluorescence microscopy showed that this loss of ER store [Ca(2+)] led to translocation of the ER-resident transmembrane Ca(2+) sensor STIM1. Using sensitive FRET-based sensors for cAMP in single cells, we further found that EPA caused a substantial increase in cellular cAMP concentration, a large fraction of which was dependent on the drop in ER [Ca(2+)], but independent of cytosolic Ca(2+). An additional component of the EPA-induced cAMP signal was sensitive to the phosphodiesterase inhibitor isobutyl methylxanthine. We conclude that EPA slowly releases ER Ca(2+) stores, resulting in the generation of cAMP. The elevated cAMP is apparently independent of classical G protein-coupled receptor activation and is likely the consequence of a newly described "store-operated" cAMP signaling pathway that is mediated by STIM1.
Figures
Similar articles
-
"Store-operated" cAMP signaling contributes to Ca2+-activated Cl- secretion in T84 colonic cells.Am J Physiol Gastrointest Liver Physiol. 2015 Oct 15;309(8):G670-9. doi: 10.1152/ajpgi.00214.2015. Epub 2015 Aug 27. Am J Physiol Gastrointest Liver Physiol. 2015. PMID: 26316590 Free PMC article.
-
Store-operated cyclic AMP signalling mediated by STIM1.Nat Cell Biol. 2009 Apr;11(4):433-42. doi: 10.1038/ncb1850. Epub 2009 Mar 15. Nat Cell Biol. 2009. PMID: 19287379
-
Termination and activation of store-operated cyclic AMP production.J Cell Mol Med. 2012 Nov;16(11):2715-25. doi: 10.1111/j.1582-4934.2012.01592.x. J Cell Mol Med. 2012. PMID: 22681560 Free PMC article.
-
Molecular physiology and pathophysiology of stromal interaction molecules.Exp Biol Med (Maywood). 2018 Mar;243(5):451-472. doi: 10.1177/1535370218754524. Epub 2018 Jan 24. Exp Biol Med (Maywood). 2018. PMID: 29363328 Free PMC article. Review.
-
Key components of store-operated Ca2+ entry in non-excitable cells.J Pharmacol Sci. 2014;125(4):340-6. doi: 10.1254/jphs.14r06cp. Epub 2014 Jul 17. J Pharmacol Sci. 2014. PMID: 25030742 Review.
Cited by
-
"Store-operated" cAMP signaling contributes to Ca2+-activated Cl- secretion in T84 colonic cells.Am J Physiol Gastrointest Liver Physiol. 2015 Oct 15;309(8):G670-9. doi: 10.1152/ajpgi.00214.2015. Epub 2015 Aug 27. Am J Physiol Gastrointest Liver Physiol. 2015. PMID: 26316590 Free PMC article.
-
Regulation by Ca2+-signaling pathways of adenylyl cyclases.Cold Spring Harb Perspect Biol. 2011 Jan 1;3(1):a004143. doi: 10.1101/cshperspect.a004143. Cold Spring Harb Perspect Biol. 2011. PMID: 21123395 Free PMC article. Review.
-
STIM1 as a key regulator for Ca2+ homeostasis in skeletal-muscle development and function.Skelet Muscle. 2011 Apr 4;1(1):16. doi: 10.1186/2044-5040-1-16. Skelet Muscle. 2011. PMID: 21798093 Free PMC article.
-
Delphinidin Reduces Glucose Uptake in Mice Jejunal Tissue and Human Intestinal Cells Lines through FFA1/GPR40.Int J Mol Sci. 2017 Apr 5;18(4):750. doi: 10.3390/ijms18040750. Int J Mol Sci. 2017. PMID: 28379159 Free PMC article.
-
Altered store operated calcium entry increases cyclic 3',5'-adenosine monophosphate production and extracellular signal-regulated kinases 1 and 2 phosphorylation in polycystin-2-defective cholangiocytes.Hepatology. 2012 Mar;55(3):856-68. doi: 10.1002/hep.24723. Hepatology. 2012. Retraction in: Hepatology. 2023 Sep 1;78(3):E65. doi: 10.1097/HEP.0000000000000484. PMID: 21987453 Free PMC article. Retracted.
References
-
- Ait-Said F, Elalamy I, Werts C, Gomard MT, Jacquemin C, Couetil JP, Hatmi M. Inhibition by eicosapentaenoic acid of IL-1beta-induced PGHS-2 expression in human microvascular endothelial cells: involvement of lipoxygenase-derived metabolites and p38 MAPK pathway. Biochim Biophys Acta 1631: 77–84, 2003 - PubMed
-
- Anti M, Armelao F, Marra G, Percesepe A, Bartoli GM, Palozza P, Parrella P, Canetta C, Gentiloni N, De Vitis I. Effects of different doses of fish oil on rectal cell proliferation in patients with sporadic colonic adenomas. Gastroenterology 107: 1709–1718, 1994 - PubMed
-
- Anti M, Marra G, Armelao F, Bartoli GM, Ficarelli R, Percesepe A, De Vitis I, Maria G, Sofo L, Rapaccini GL. Effect of omega-3 fatty acids on rectal mucosal cell proliferation in subjects at risk for colon cancer. Gastroenterology 103: 883–891, 1992 - PubMed
-
- Bartram HP, Gostner A, Scheppach W, Reddy BS, Rao CV, Dusel G, Richter F, Richter A, Kasper H. Effects of fish oil on rectal cell proliferation, mucosal fatty acids, and prostaglandin E2 release in healthy subjects. Gastroenterology 105: 1317–1322, 1993 - PubMed
-
- Belluzzi A, Boschi S, Brignola C, Munarini A, Cariani G, Miglio F. Polyunsaturated fatty acids and inflammatory bowel disease. Am J Clin Nutr 71: 339S–342S, 2000 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous