

Adrenal glucocorticoids have a key role in circadian resynchronization in a mouse model of jet lag
- PMID: 20577050
- PMCID: PMC2898589
- DOI: 10.1172/JCI41192
Adrenal glucocorticoids have a key role in circadian resynchronization in a mouse model of jet lag
Abstract
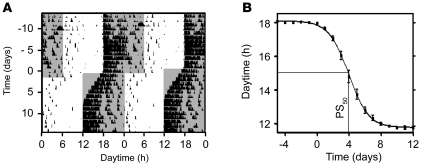
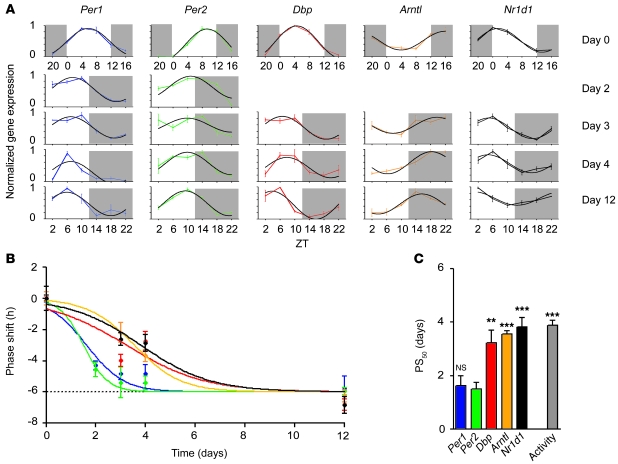
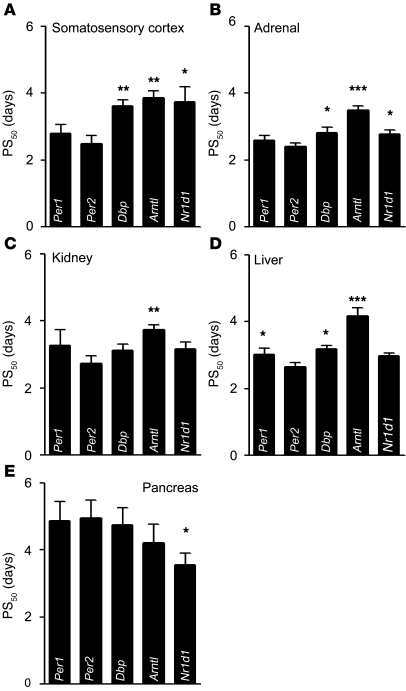
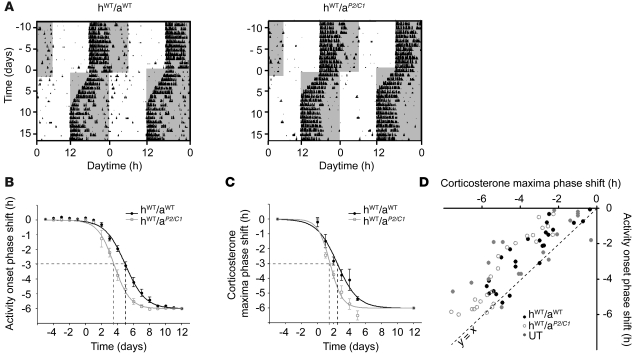
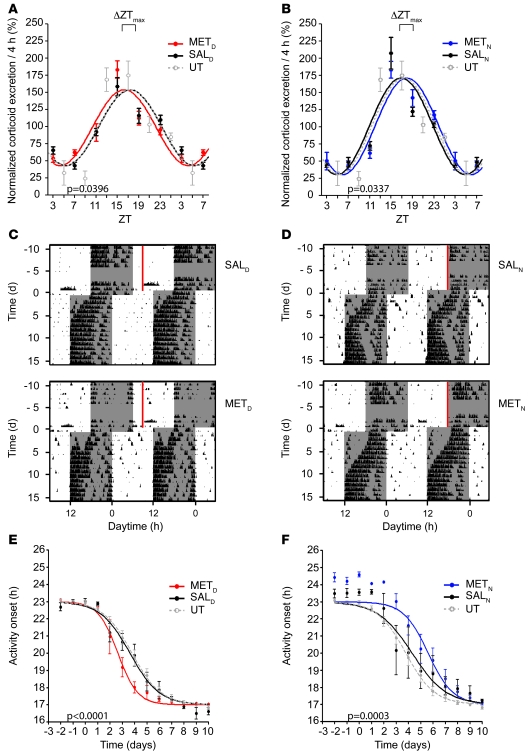
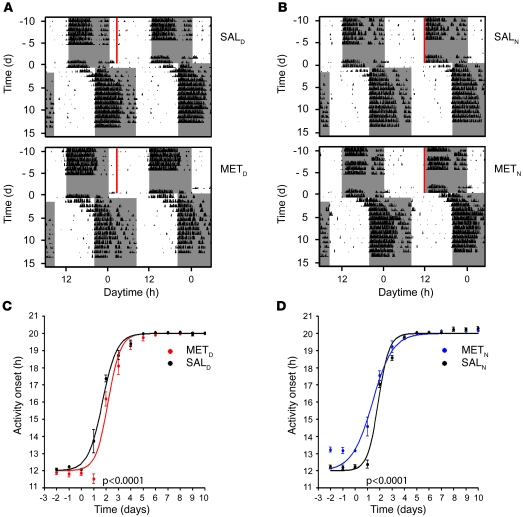
Jet lag encompasses a range of psycho- and physiopathological symptoms that arise from temporal misalignment of the endogenous circadian clock with external time. Repeated jet lag exposure, encountered by business travelers and airline personnel as well as shift workers, has been correlated with immune deficiency, mood disorders, elevated cancer risk, and anatomical anomalies of the forebrain. Here, we have characterized the molecular response of the mouse circadian system in an established experimental paradigm for jet lag whereby mice entrained to a 12-hour light/12-hour dark cycle undergo light phase advancement by 6 hours. Unexpectedly, strong heterogeneity of entrainment kinetics was found not only between different organs, but also within the molecular clockwork of each tissue. Manipulation of the adrenal circadian clock, in particular phase-shifting of adrenal glucocorticoid rhythms, regulated the speed of behavioral reentrainment. Blocking adrenal corticosterone either prolonged or shortened jet lag, depending on the time of administration. This key role of adrenal glucocorticoid phasing for resetting of the circadian system provides what we believe to be a novel mechanism-based approach for possible therapies for jet lag and jet lag-associated diseases.
Figures
Comment in
-
Location, location, location: important for jet-lagged circadian loops.J Clin Invest. 2010 Jul;120(7):2265-7. doi: 10.1172/JCI43632. Epub 2010 Jun 23. J Clin Invest. 2010. PMID: 20577055 Free PMC article. Review.
Similar articles
-
Accelerating recovery from jet lag: prediction from a multi-oscillator model and its experimental confirmation in model animals.Sci Rep. 2017 Apr 26;7:46702. doi: 10.1038/srep46702. Sci Rep. 2017. PMID: 28443630 Free PMC article.
-
Reentrainment of the circadian pacemaker during jet lag: East-west asymmetry and the effects of north-south travel.J Theor Biol. 2018 Jan 21;437:261-285. doi: 10.1016/j.jtbi.2017.10.002. Epub 2017 Oct 4. J Theor Biol. 2018. PMID: 28987464
-
Effects of chronic jet lag on the central and peripheral circadian clocks in CBA/N mice.Chronobiol Int. 2014 Mar;31(2):189-98. doi: 10.3109/07420528.2013.837478. Epub 2013 Oct 22. Chronobiol Int. 2014. PMID: 24147659
-
From circadian clock mechanism to sleep disorders and jet lag: Insights from a computational approach.Biochem Pharmacol. 2021 Sep;191:114482. doi: 10.1016/j.bcp.2021.114482. Epub 2021 Feb 20. Biochem Pharmacol. 2021. PMID: 33617843 Review.
-
Circadian rhythm sleep disorders: pathophysiology and potential approaches to management.CNS Drugs. 2001;15(4):311-28. doi: 10.2165/00023210-200115040-00005. CNS Drugs. 2001. PMID: 11463135 Review.
Cited by
-
Inducible cAMP early repressor regulates the Period 1 gene of the hepatic and adrenal clocks.J Biol Chem. 2013 Apr 12;288(15):10318-27. doi: 10.1074/jbc.M112.445692. Epub 2013 Feb 25. J Biol Chem. 2013. PMID: 23443664 Free PMC article.
-
Circadian disruption alters mouse lung clock gene expression and lung mechanics.J Appl Physiol (1985). 2012 Aug;113(3):385-92. doi: 10.1152/japplphysiol.00244.2012. Epub 2012 Jun 7. J Appl Physiol (1985). 2012. PMID: 22678966 Free PMC article.
-
Deregulated Brain's Central Clock Management on Sleep-Wake Behavior in Women With Polycystic Ovary Syndrome: Melatonin & Sleep Pattern.J Family Reprod Health. 2022 Dec;16(4):229-238. doi: 10.18502/jfrh.v16i4.11348. J Family Reprod Health. 2022. PMID: 37465433 Free PMC article. Review.
-
Light stimulates the mouse adrenal through a retinohypothalamic pathway independent of an effect on the clock in the suprachiasmatic nucleus.PLoS One. 2014 Mar 21;9(3):e92959. doi: 10.1371/journal.pone.0092959. eCollection 2014. PLoS One. 2014. PMID: 24658072 Free PMC article.
-
Vestibular stimulation by 2G hypergravity modifies resynchronization in temperature rhythm in rats.Sci Rep. 2020 Jun 8;10(1):9216. doi: 10.1038/s41598-020-65496-x. Sci Rep. 2020. PMID: 32514078 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
