

Sustained calcium signalling and caspase-3 activation involve NMDA receptors in thymocytes in contact with dendritic cells
- PMID: 20577261
- PMCID: PMC3131881
- DOI: 10.1038/cdd.2010.79
Sustained calcium signalling and caspase-3 activation involve NMDA receptors in thymocytes in contact with dendritic cells
Abstract
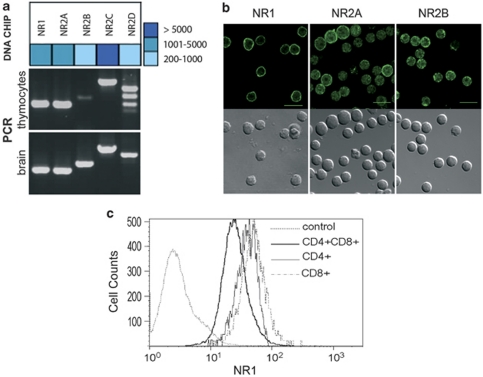
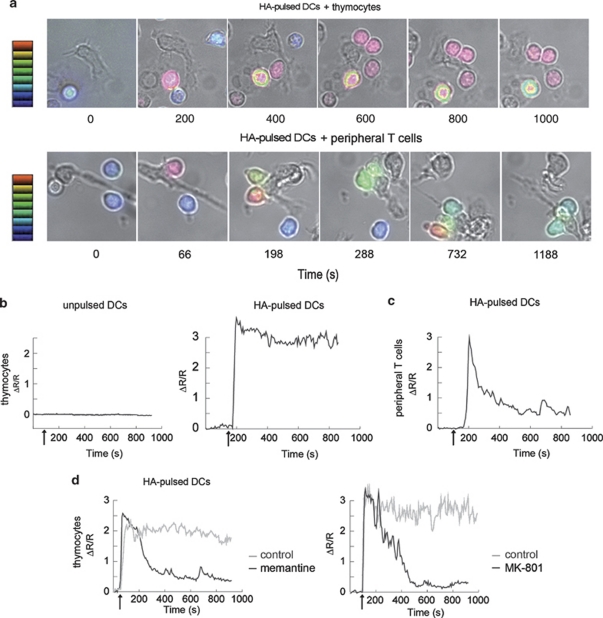
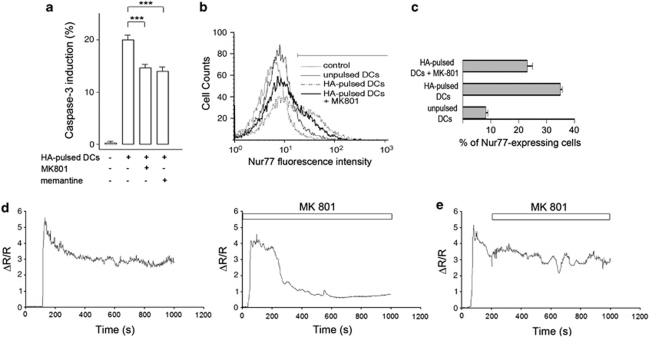
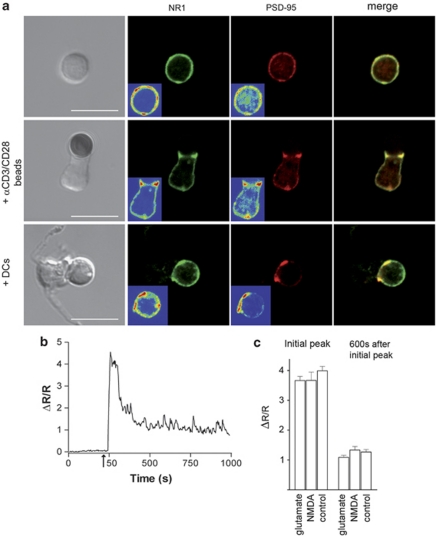
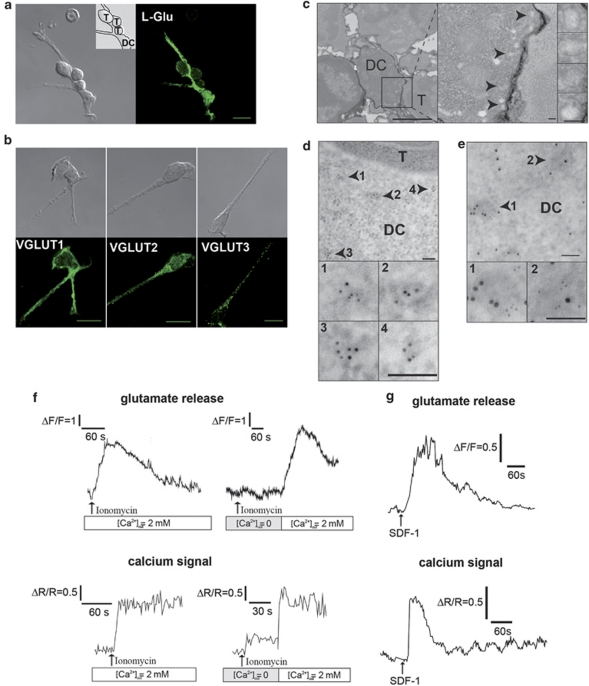
L-glutamate, the major excitatory neurotransmitter, also has a role in non-neuronal tissues and modulates immune responses. Whether NMDA receptor (NMDAR) signalling is involved in T-cell development is unknown. In this study, we show that mouse thymocytes expressed an array of glutamate receptors, including NMDARs subunits. Sustained calcium (Ca(2+)) signals and caspase-3 activation in thymocytes were induced by interaction with antigen-pulsed dendritic cells (DCs) and were inhibited by NMDAR antagonists MK801 and memantine. NMDARs were transiently activated, triggered the sustained Ca(2+) signal and were corecruited with the PDZ-domain adaptor postsynaptic density (PSD)-95 to thymocyte-DC contact zones. Although T-cell receptor (TCR) activation was sufficient for relocalization of NMDAR and PSD-95 at the contact zone, NMDAR could be activated only in a synaptic context. In these T-DC contacts, thymocyte activation occurred in the absence of exogenous glutamate, indicating that DCs could be a physiological source of glutamate. DCs expressed glutamate, glutamate-specific vesicular glutamate transporters and were capable of fast glutamate release through a Ca(2+)-dependent mechanism. We suggest that glutamate released by DCs could elicit focal responses through NMDAR-signalling in T cells undergoing apoptosis. Thus, synapses between T and DCs could provide a functional platform for coupling TCR activation and NMDAR signalling, which might reflect on T-cell development and modulation of the immune response.
Figures
References
-
- Nedergaard M, Takano T, Hansen AJ. Beyond the role of glutamate as a neurotransmitter. Nat Rev Neurosci. 2002;3:748–755. - PubMed
-
- Skerry TM, Genever PG. Glutamate signalling in non-neuronal tissues. Trends Pharmacol Sci. 2001;22:174–181. - PubMed
-
- Pacheco R, Gallart T, Lluis C, Franco R. Role of glutamate on T-cell mediated immunity. J Neuroimmunol. 2007;185:9–19. - PubMed
-
- Pacheco R, Ciruela F, Casado V, Mallol J, Gallart T, Lluis C, et al. Group I metabotropic glutamate receptors mediate a dual role of glutamate in T cell activation. J Biol Chem. 2004;279:33352–33358. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
