

Toxoplasma gondii activates hypoxia-inducible factor (HIF) by stabilizing the HIF-1alpha subunit via type I activin-like receptor kinase receptor signaling
- PMID: 20581113
- PMCID: PMC2930684
- DOI: 10.1074/jbc.M110.147041
Toxoplasma gondii activates hypoxia-inducible factor (HIF) by stabilizing the HIF-1alpha subunit via type I activin-like receptor kinase receptor signaling
Abstract
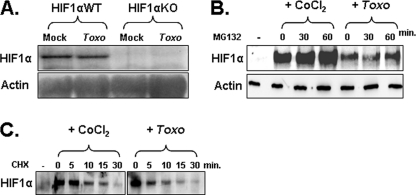
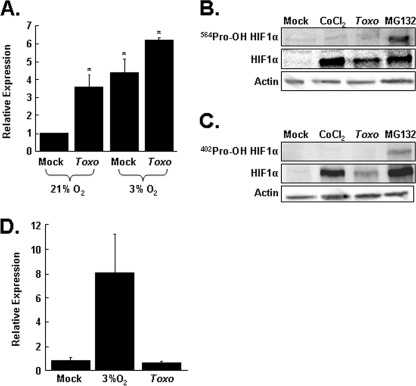
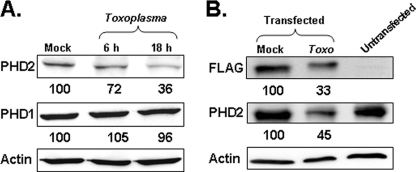
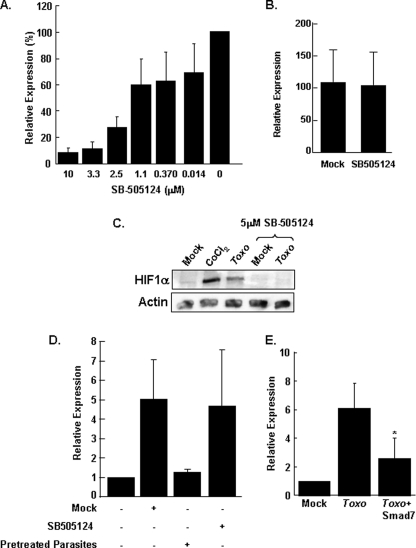
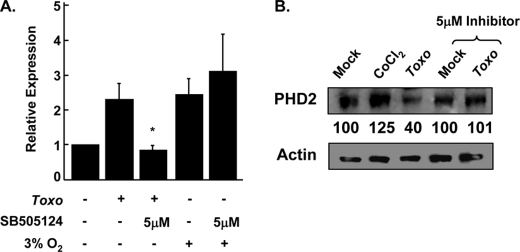
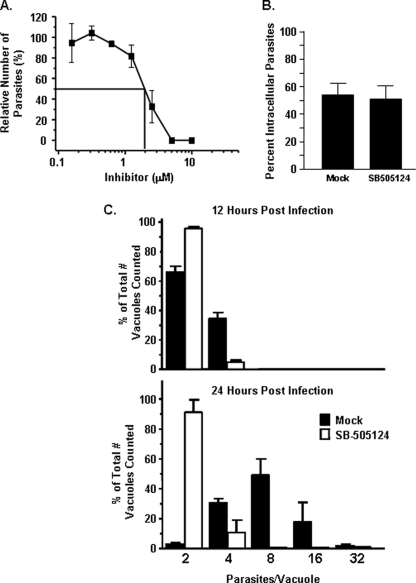
Toxoplasma gondii is an intracellular protozoan parasite that can cause devastating disease in fetuses and immune-compromised individuals. We previously reported that the alpha subunit of the host cell transcription factor, hypoxia-inducible factor-1 (HIF-1), is up-regulated by infection and necessary for Toxoplasma growth. Under basal conditions, HIF-1alpha is constitutively expressed but rapidly targeted for proteasomal degradation after two proline residues are hydroxylated by a family of prolyl hydroxylases (PHDs). The PHDs are alpha-ketoglutarate-dependent dioxygenases that have low K(m) values for oxygen, making them important cellular oxygen sensors. Thus, when oxygen levels decrease, HIF-1alpha is not hydroxylated, and HIF-1 is activated. How Toxoplasma activates HIF-1 under normoxic conditions remains unknown. Here, we report that Toxoplasma infection increases HIF-1alpha stability by preventing HIF-1alpha prolyl hydroxylation. Infection significantly decreases PHD2 abundance, which is the key prolyl hydroxylase for regulating HIF-1alpha. The effects of Toxoplasma on HIF-1alpha abundance and prolyl hydroxylase activity require activin-like receptor kinase signaling. Finally, parasite growth is severely diminished when signaling from this family of receptors is inhibited. Together, these data indicate that PHD2 is a key host cell factor for T. gondii growth and represent a novel mechanism by which a microbial pathogen subverts host cell signaling and transcription to establish its replicative niche.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
