

The pre-mRNA splicing machinery of trypanosomes: complex or simplified?
- PMID: 20581293
- PMCID: PMC2918933
- DOI: 10.1128/EC.00113-10
The pre-mRNA splicing machinery of trypanosomes: complex or simplified?
Abstract
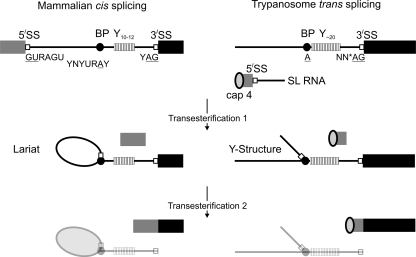
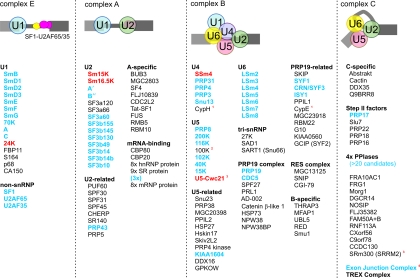
Trypanosomatids are early-diverged, protistan parasites of which Trypanosoma brucei, Trypanosoma cruzi, and several species of Leishmania cause severe, often lethal diseases in humans. To better combat these parasites, their molecular biology has been a research focus for more than 3 decades, and the discovery of spliced leader (SL) trans splicing in T. brucei established a key difference between parasites and hosts. In SL trans splicing, the capped 5'-terminal region of the small nuclear SL RNA is fused onto the 5' end of each mRNA. This process, in conjunction with polyadenylation, generates individual mRNAs from polycistronic precursors and creates functional mRNA by providing the cap structure. The reaction is a two-step transesterification process analogous to intron removal by cis splicing which, in trypanosomatids, is confined to very few pre-mRNAs. Both types of pre-mRNA splicing are carried out by the spliceosome, consisting of five U-rich small nuclear RNAs (U snRNAs) and, in humans, up to approximately 170 different proteins. While trypanosomatids possess a full set of spliceosomal U snRNAs, only a few splicing factors were identified by standard genome annotation because trypanosomatid amino acid sequences are among the most divergent in the eukaryotic kingdom. This review focuses on recent progress made in the characterization of the splicing factor repertoire in T. brucei, achieved by tandem affinity purification of splicing complexes, by systematic analysis of proteins containing RNA recognition motifs, and by mining the genome database. In addition, recent findings about functional differences between trypanosome and human pre-mRNA splicing factors are discussed.
Figures
References
-
- Aslett M., Aurrecoechea C., Berriman M., Brestelli J., Brunk B. P., Carrington M., Depledge D. P., Fischer S., Gajria B., Gao X., Gardner M. J., Gingle A., Grant G., Harb O. S., Heiges M., Hertz-Fowler C., Houston R., Innamorato F., Iodice J., Kissinger J. C., Kraemer E., Li W., Logan F. J., Miller J. A., Mitra S., Myler P. J., Nayak V., Pennington C., Phan I., Pinney D. F., Ramasamy G., Rogers M. B., Roos D. S., Ross C., Sivam D., Smith D. F., Srinivasamoorthy G., Stoeckert C. J., Jr., Subramanian S., Thibodeau R., Tivey A., Treatman C., Velarde G., Wang H. 2010. TriTrypDB: a functional genomic resource for the Trypanosomatidae. Nucleic Acids Res. 38:D457–D462 - PMC - PubMed
-
- Avila M. L., Bercovich N., Westergaard G., Levin M. J., Vazquez M. P. 2007. Mapping of the protein-binding interface between splicing factors SF3b155 and p14 of Trypanosoma cruzi. Biochem. Biophys. Res. Commun. 364:26–32 - PubMed
-
- Bangs J. D., Crain P. F., Hashizume T., McCloskey J. A., Boothroyd J. C. 1992. Mass spectrometry of mRNA cap 4 from trypanosomatids reveals two novel nucleosides. J. Biol. Chem. 267:9805–9815 - PubMed
-
- Barrandon C., Spiluttini B., Bensaude O. 2008. Non-coding RNAs regulating the transcriptional machinery. Biol. Cell 100:83–95 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
