

Complement receptor 1 is a sialic acid-independent erythrocyte receptor of Plasmodium falciparum
- PMID: 20585558
- PMCID: PMC2887475
- DOI: 10.1371/journal.ppat.1000968
Complement receptor 1 is a sialic acid-independent erythrocyte receptor of Plasmodium falciparum
Abstract
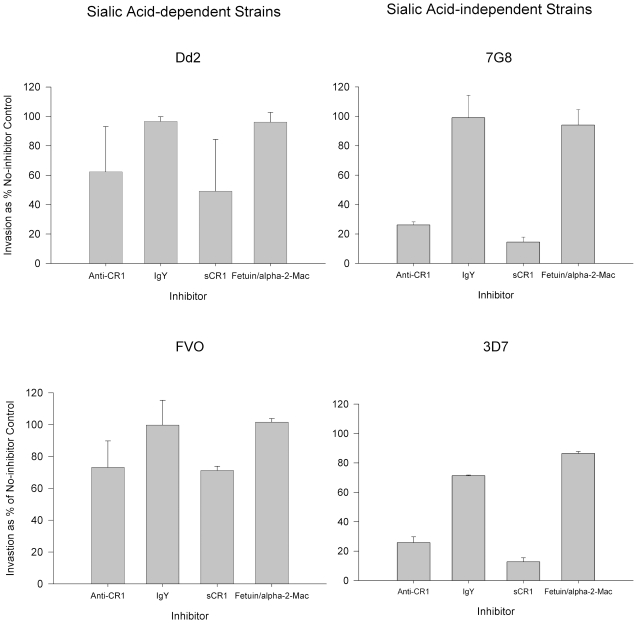
Plasmodium falciparum is a highly lethal malaria parasite of humans. A major portion of its life cycle is dedicated to invading and multiplying inside erythrocytes. The molecular mechanisms of erythrocyte invasion are incompletely understood. P. falciparum depends heavily on sialic acid present on glycophorins to invade erythrocytes. However, a significant proportion of laboratory and field isolates are also able to invade erythrocytes in a sialic acid-independent manner. The identity of the erythrocyte sialic acid-independent receptor has been a mystery for decades. We report here that the complement receptor 1 (CR1) is a sialic acid-independent receptor for the invasion of erythrocytes by P. falciparum. We show that soluble CR1 (sCR1) as well as polyclonal and monoclonal antibodies against CR1 inhibit sialic acid-independent invasion in a variety of laboratory strains and wild isolates, and that merozoites interact directly with CR1 on the erythrocyte surface and with sCR1-coated microspheres. Also, the invasion of neuraminidase-treated erythrocytes correlates with the level of CR1 expression. Finally, both sialic acid-independent and dependent strains invade CR1 transgenic mouse erythrocytes preferentially over wild-type erythrocytes but invasion by the latter is more sensitive to neuraminidase. These results suggest that both sialic acid-dependent and independent strains interact with CR1 in the normal red cell during the invasion process. However, only sialic acid-independent strains can do so without the presence of glycophorin sialic acid. Our results close a longstanding and important gap in the understanding of the mechanism of erythrocyte invasion by P. falciparum that will eventually make possible the development of an effective blood stage vaccine.
Conflict of interest statement
The authors have declared that no competing interests exist.
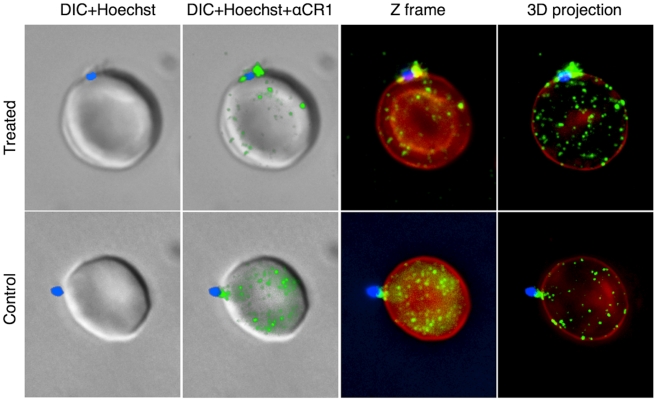
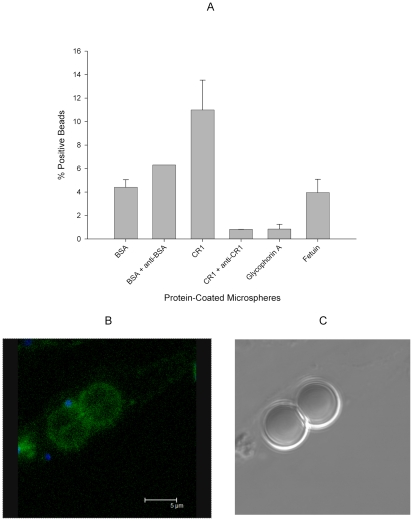
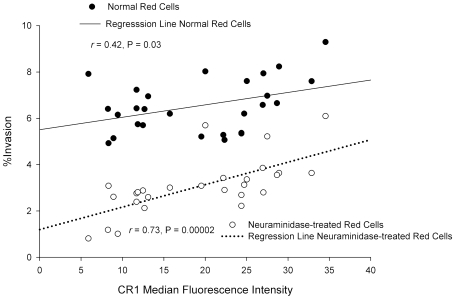
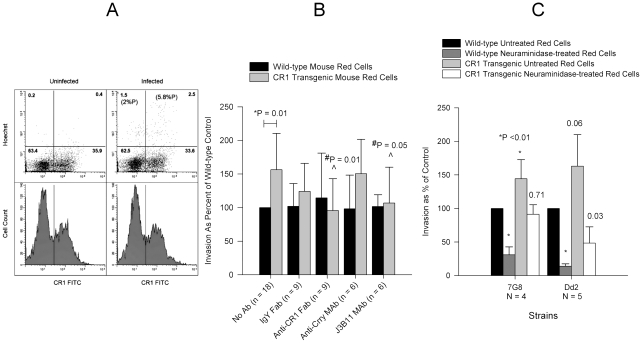
Figures
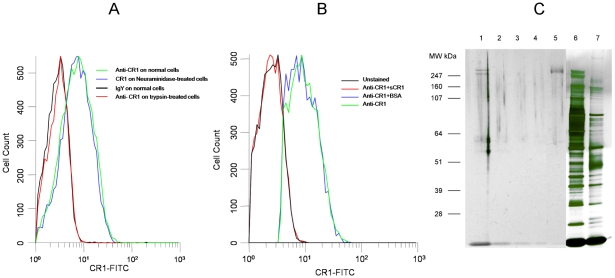
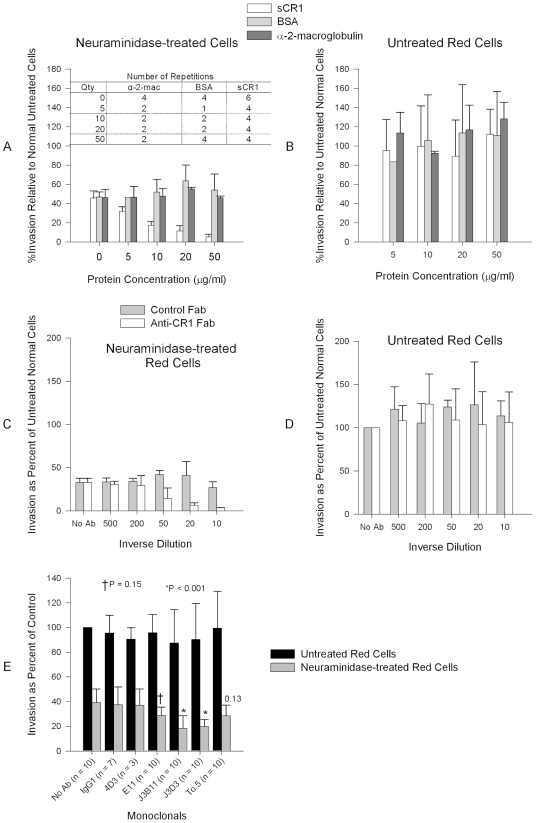
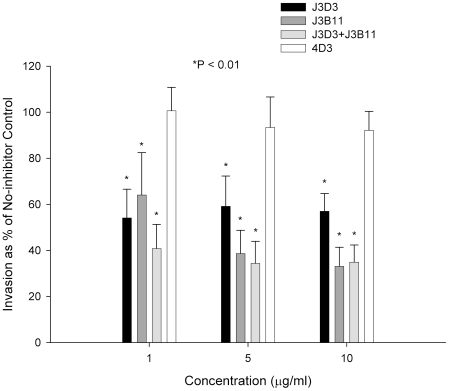
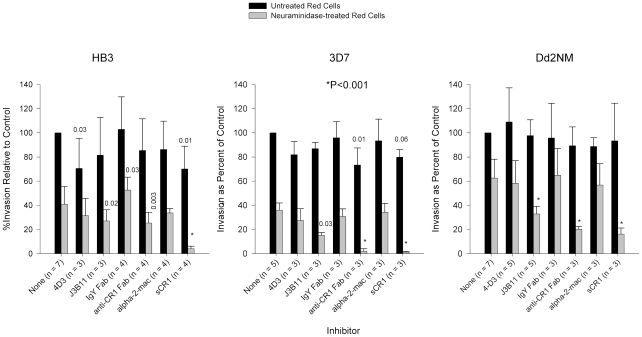
References
-
- Deas JE, Lee LT. Competitive inhibition by soluble erythrocyte glycoproteins of penetration by Plasmodium falciparum. Am J Trop Med Hyg. 1981;30:1164–1167. - PubMed
-
- Pasvol G, Jungery M, Weatherall DJ, Parsons SF, Anstee DJ, et al. Glycophorin as a possible receptor for Plasmodium falciparum. Lancet. 1982;2:947–950. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
